

Gradual emergence of song selectivity in sensorimotor structures of the male zebra finch song system
- PMID: 10366643
- PMCID: PMC6782639
- DOI: 10.1523/JNEUROSCI.19-12-05108.1999
Gradual emergence of song selectivity in sensorimotor structures of the male zebra finch song system
Abstract
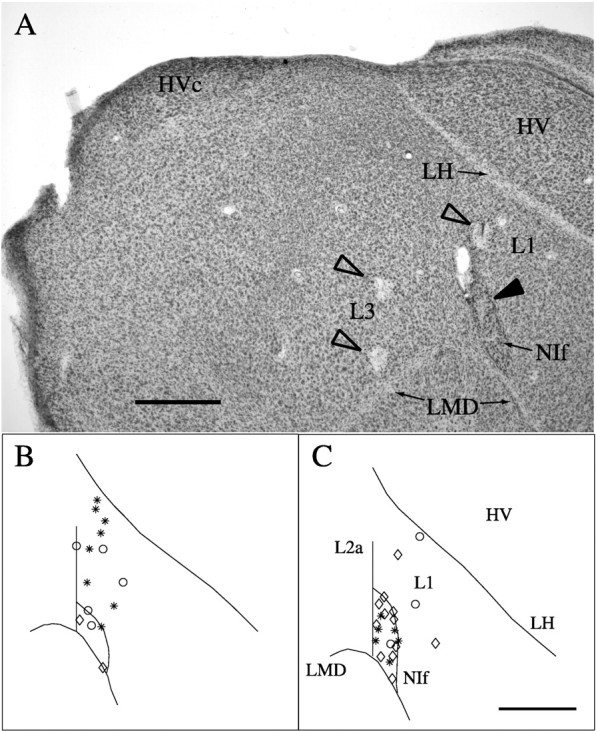
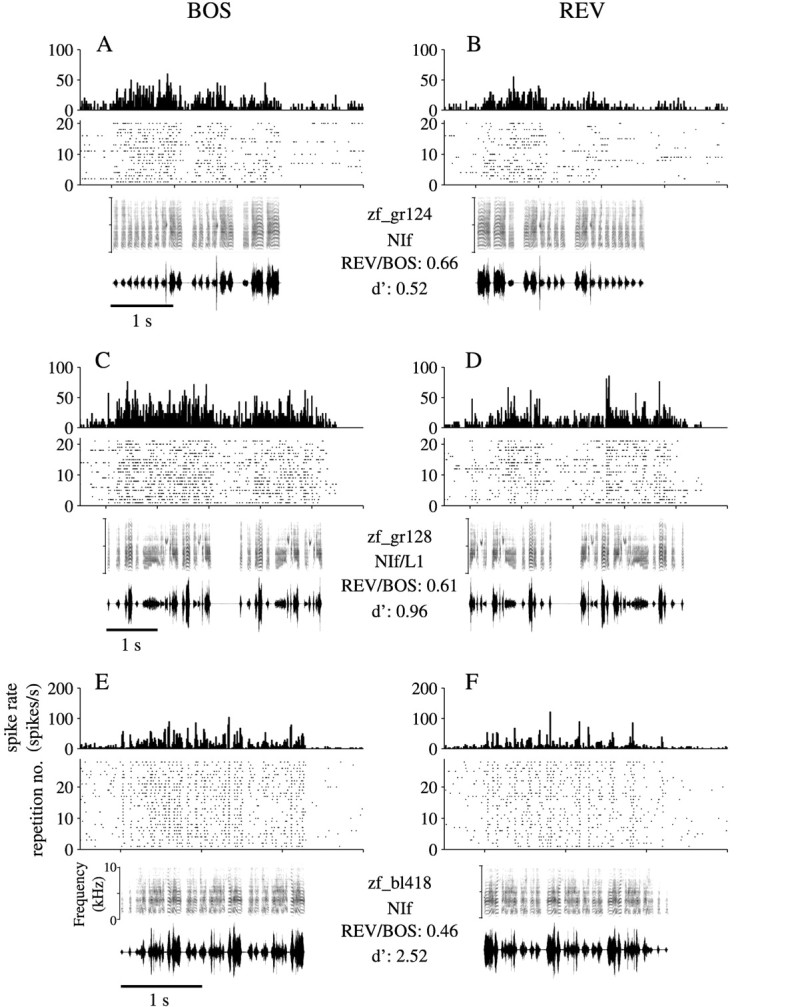
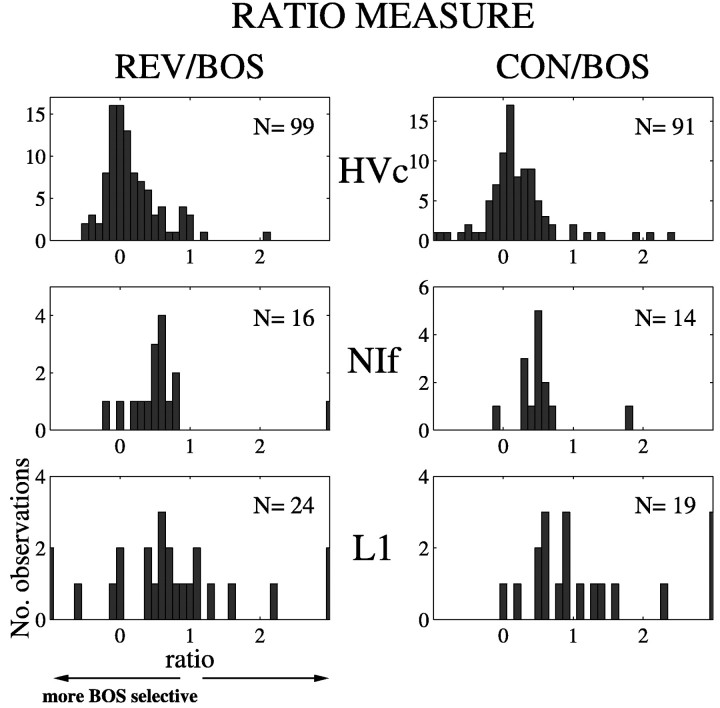
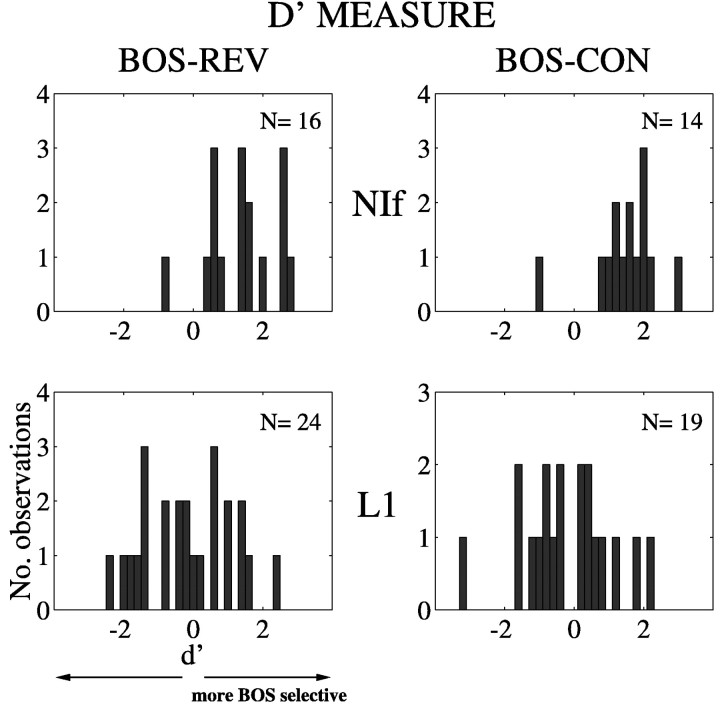
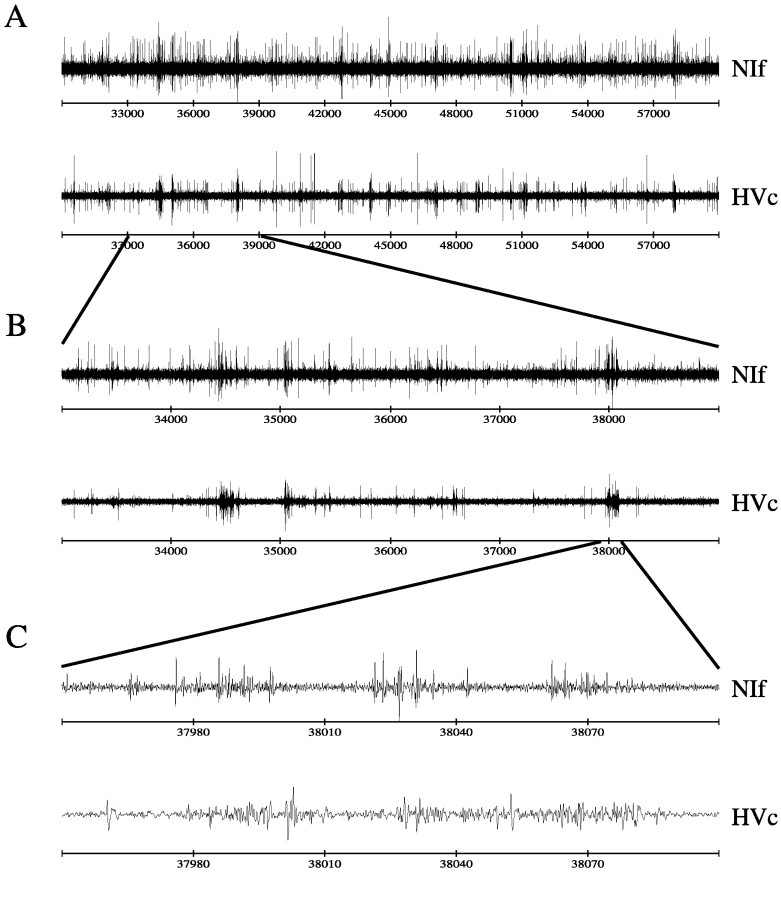
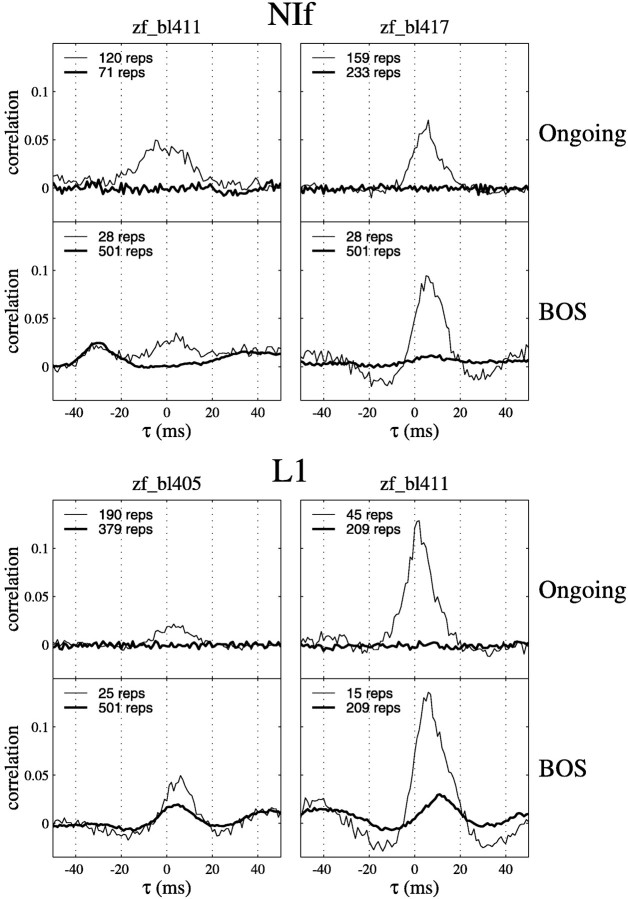
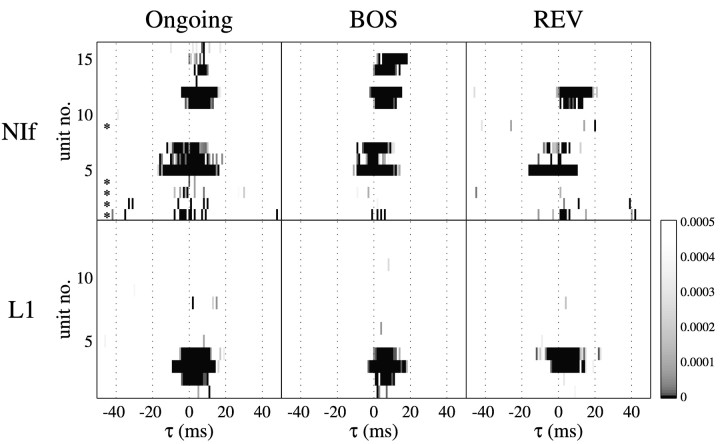
Birdsong is a model system for understanding how motor and sensory information interact to coordinate behavior. Neurons in one potential site of sensorimotor integration, the forebrain nucleus HVc, have premotor activity during singing and auditory activity during playback of the bird's own song. It is not known whether the high degree of selectivity for learned features of song observed during playback arises in HVc or also in structures afferent to HVc. We recorded in anesthetized adult zebra finches from two structures afferent to HVc: either the nucleus interfacialis (NIf) or the L1 subdivision of the field L complex, and simultaneously from a second electrode in HVc. Correlations in the bursting pattern of ongoing activity of HVc and NIf recordings were observed; these helped to localize the first electrode to NIf recording sites. Most NIf neurons exhibited song-selective responses, but as a population, they were less selective than were HVc neurons. Most L1 neurons were not song-selective. NIf neurons have also been reported to have premotor activity during singing; thus, NIf is another potential site of auditory-motor interactions in the song system. Evidence gathered to date suggests that those brain areas in the passerine forebrain that are recruited during song production also display the most selective learned auditory responses.
Figures
References
-
- Bedenbaugh P, Gerstein GL. Multiunit normalized cross correlation differs from the average single-unit normalized correlation. Neural Comput. 1997;9:1265–1275. - PubMed
-
- Capsius B, Leppelsack HJ. Influence of urethane anesthesia on neural processing in the auditory cortex analogue of a songbird. Hear Res. 1996;96:59–70. - PubMed
-
- Eales LA. Song learning in zebra finches: some effects of song model availability on what is learnt and when. Anim Behav. 1985;33:1293–1300.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources