

The glucuronic acid utilization gene cluster from Bacillus stearothermophilus T-6
- PMID: 10368143
- PMCID: PMC93846
- DOI: 10.1128/JB.181.12.3695-3704.1999
The glucuronic acid utilization gene cluster from Bacillus stearothermophilus T-6
Abstract
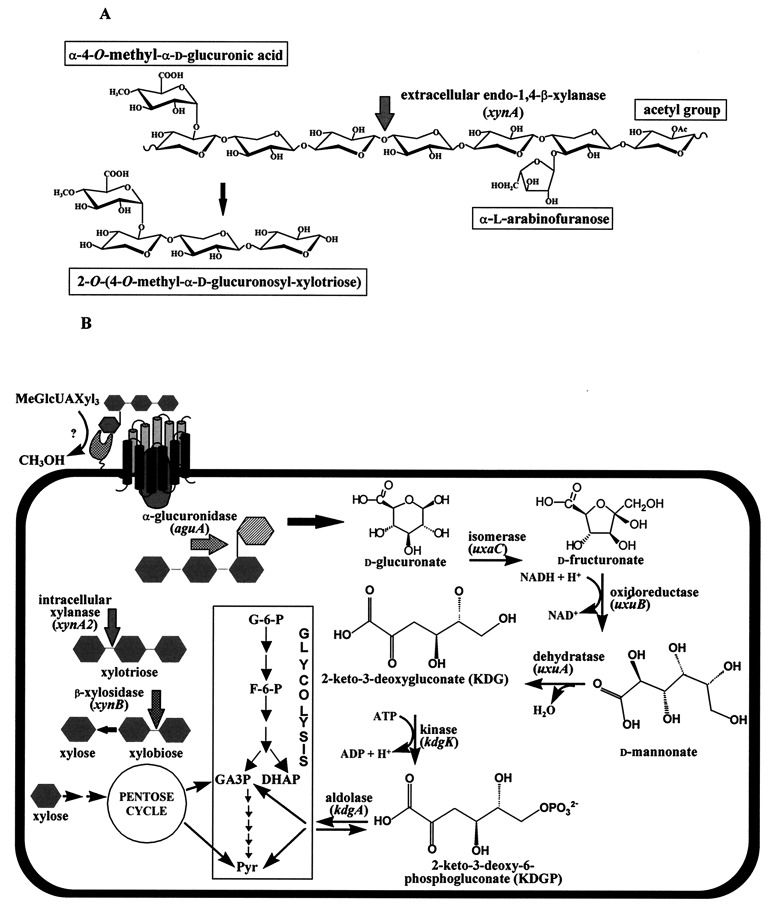
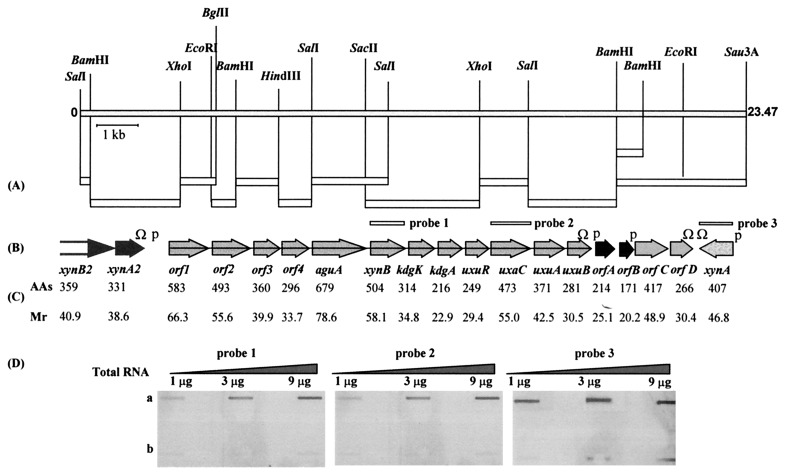
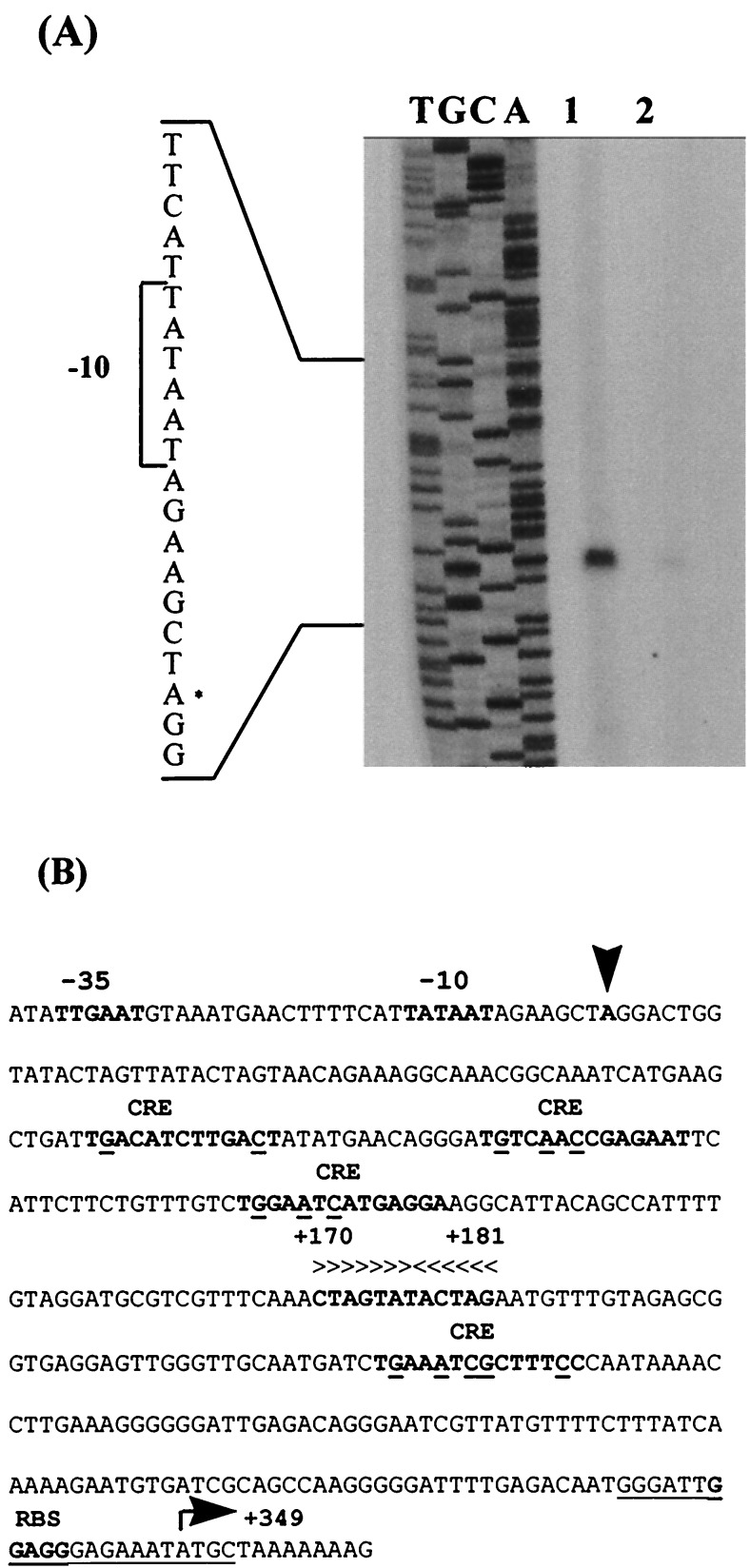
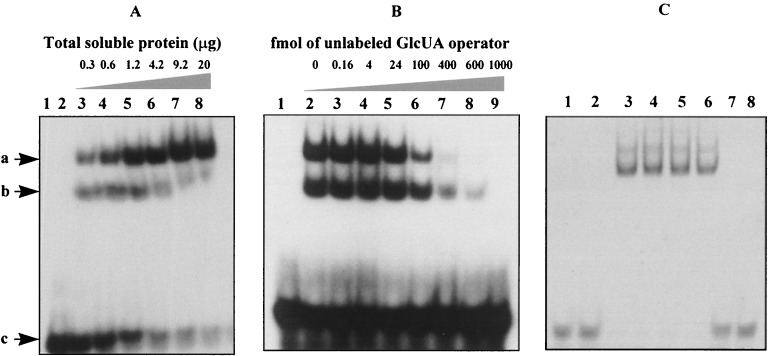
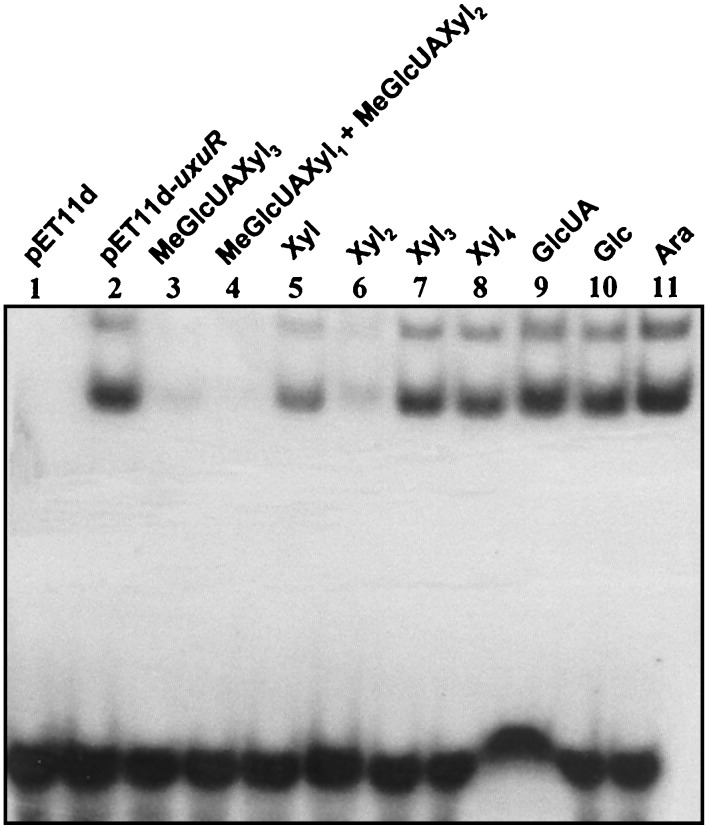
A lambda-EMBL3 genomic library of Bacillus stearothermophilus T-6 was screened for hemicellulolytic activities, and five independent clones exhibiting beta-xylosidase activity were isolated. The clones overlap each other and together represent a 23.5-kb chromosomal segment. The segment contains a cluster of xylan utilization genes, which are organized in at least three transcriptional units. These include the gene for the extracellular xylanase, xylanase T-6; part of an operon coding for an intracellular xylanase and a beta-xylosidase; and a putative 15.5-kb-long transcriptional unit, consisting of 12 genes involved in the utilization of alpha-D-glucuronic acid (GlcUA). The first four genes in the potential GlcUA operon (orf1, -2, -3, and -4) code for a putative sugar transport system with characteristic components of the binding-protein-dependent transport systems. The most likely natural substrate for this transport system is aldotetraouronic acid [2-O-alpha-(4-O-methyl-alpha-D-glucuronosyl)-xylotriose] (MeGlcUAXyl3). The following two genes code for an intracellular alpha-glucuronidase (aguA) and a beta-xylosidase (xynB). Five more genes (kdgK, kdgA, uxaC, uxuA, and uxuB) encode proteins that are homologous to enzymes involved in galacturonate and glucuronate catabolism. The gene cluster also includes a potential regulatory gene, uxuR, the product of which resembles repressors of the GntR family. The apparent transcriptional start point of the cluster was determined by primer extension analysis and is located 349 bp from the initial ATG codon. The potential operator site is a perfect 12-bp inverted repeat located downstream from the promoter between nucleotides +170 and +181. Gel retardation assays indicated that UxuR binds specifically to this sequence and that this binding is efficiently prevented in vitro by MeGlcUAXyl3, the most likely molecular inducer.
Figures
Similar articles
-
Identification and characterization of clustered genes for thermostable xylan-degrading enzymes, beta-xylosidase and xylanase, of Bacillus stearothermophilus 21.Appl Environ Microbiol. 1994 Jul;60(7):2252-8. doi: 10.1128/aem.60.7.2252-2258.1994. Appl Environ Microbiol. 1994. PMID: 8074507 Free PMC article.
-
Molecular characterization of a high-affinity xylobiose transporter of Streptomyces thermoviolaceus OPC-520 and its transcriptional regulation.J Bacteriol. 2004 Feb;186(4):1029-37. doi: 10.1128/JB.186.4.1029-1037.2004. J Bacteriol. 2004. PMID: 14761997 Free PMC article.
-
Hexuronate catabolism in Erwinia chrysanthemi.J Bacteriol. 1987 Mar;169(3):1223-31. doi: 10.1128/jb.169.3.1223-1231.1987. J Bacteriol. 1987. PMID: 3029026 Free PMC article.
-
Cloning and sequencing of a cellobiose phosphotransferase system operon from Bacillus stearothermophilus XL-65-6 and functional expression in Escherichia coli.J Bacteriol. 1993 Oct;175(20):6441-50. doi: 10.1128/jb.175.20.6441-6450.1993. J Bacteriol. 1993. PMID: 8407820 Free PMC article.
-
The L-Arabinan utilization system of Geobacillus stearothermophilus.J Bacteriol. 2011 Jun;193(11):2838-50. doi: 10.1128/JB.00222-11. Epub 2011 Apr 1. J Bacteriol. 2011. PMID: 21460081 Free PMC article.
Cited by
-
Effect of dimer dissociation on activity and thermostability of the alpha-glucuronidase from Geobacillus stearothermophilus: dissecting the different oligomeric forms of family 67 glycoside hydrolases.J Bacteriol. 2004 Oct;186(20):6928-37. doi: 10.1128/JB.186.20.6928-6937.2004. J Bacteriol. 2004. PMID: 15466046 Free PMC article.
-
Complete genome sequences of Geobacillus sp. Y412MC52, a xylan-degrading strain isolated from obsidian hot spring in Yellowstone National Park.Stand Genomic Sci. 2015 Oct 19;10:81. doi: 10.1186/s40793-015-0075-0. eCollection 2015. Stand Genomic Sci. 2015. PMID: 26500717 Free PMC article.
-
Reconstitution of a thermostable xylan-degrading enzyme mixture from the bacterium Caldicellulosiruptor bescii.Appl Environ Microbiol. 2013 Mar;79(5):1481-90. doi: 10.1128/AEM.03265-12. Epub 2012 Dec 21. Appl Environ Microbiol. 2013. PMID: 23263957 Free PMC article.
-
Dimethyl adenosine transferase (KsgA) deficiency in Salmonella enterica Serovar Enteritidis confers susceptibility to high osmolarity and virulence attenuation in chickens.Appl Environ Microbiol. 2013 Dec;79(24):7857-66. doi: 10.1128/AEM.03040-13. Epub 2013 Oct 11. Appl Environ Microbiol. 2013. PMID: 24123731 Free PMC article.
-
Preliminary crystallographic analysis of Xyn52B2, a GH52 β-D-xylosidase from Geobacillus stearothermophilus T6.Acta Crystallogr F Struct Biol Commun. 2014 Dec 1;70(Pt 12):1675-82. doi: 10.1107/S2053230X14023887. Epub 2014 Nov 28. Acta Crystallogr F Struct Biol Commun. 2014. PMID: 25484225 Free PMC article.
References
-
- Adams M D, Wagner L M, Graddis T J, Landick R, Antonucci T K, Gibson A L, Oxender D L. Nucleotide sequence and genetic characterization reveal six essential genes for the LIV-I and LS transport system of Escherichia coli. J Biol Chem. 1990;265:11436–11443. - PubMed
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Ames G F-L. Bacterial periplasmic transport systems: structure, mechanism and evolution. Annu Rev Biochem. 1986;55:397–425. - PubMed
-
- Ames G F-L, Mimura C S, Shyamala V. Bacterial periplasmic permeases belong to a family of transport proteins operating from Escherichia colito human: traffic ATPases. FEMS Microbiol Rev. 1990;75:429–446. - PubMed
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J G, Smith J A, Struhl K. Short protocols in molecular biology. New York, N.Y: John Wiley & Sons, Inc.; 1992.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
