

Bacterioferritin A modulates catalase A (KatA) activity and resistance to hydrogen peroxide in Pseudomonas aeruginosa
- PMID: 10368148
- PMCID: PMC93851
- DOI: 10.1128/JB.181.12.3730-3742.1999
Bacterioferritin A modulates catalase A (KatA) activity and resistance to hydrogen peroxide in Pseudomonas aeruginosa
Abstract
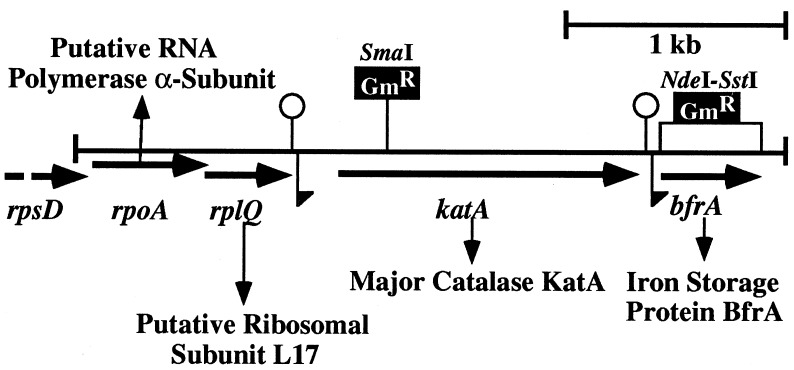
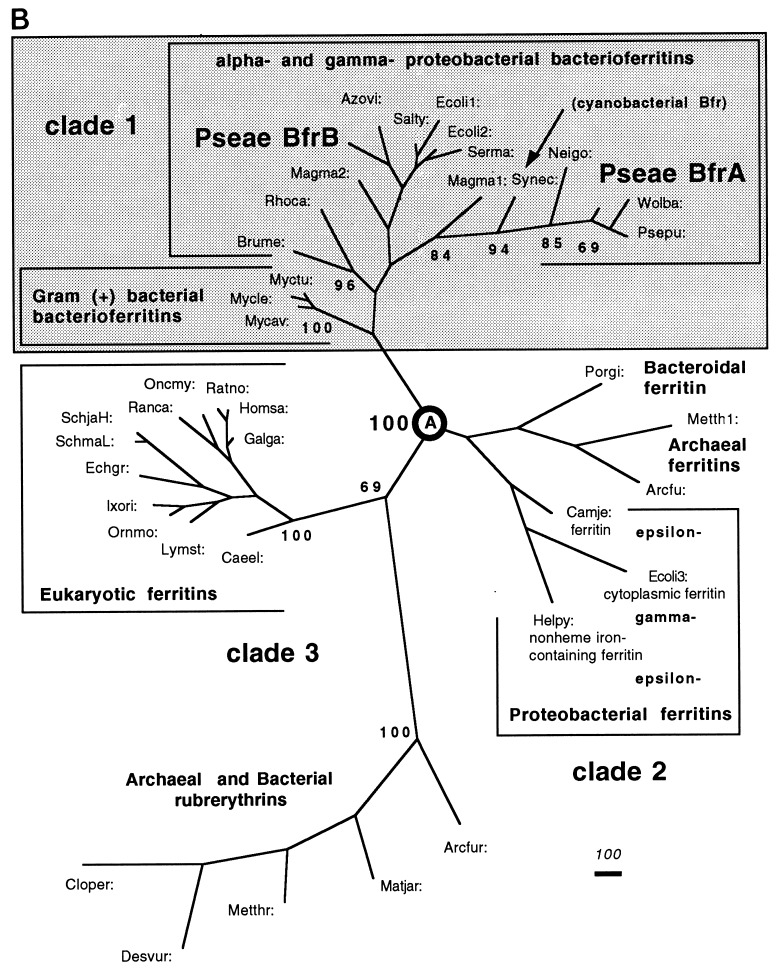
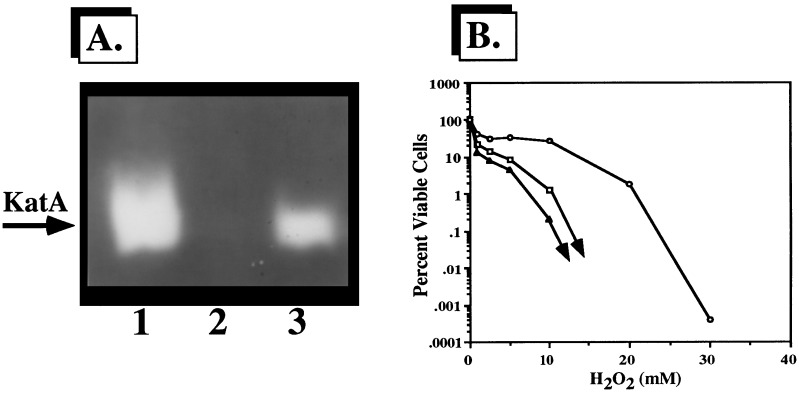
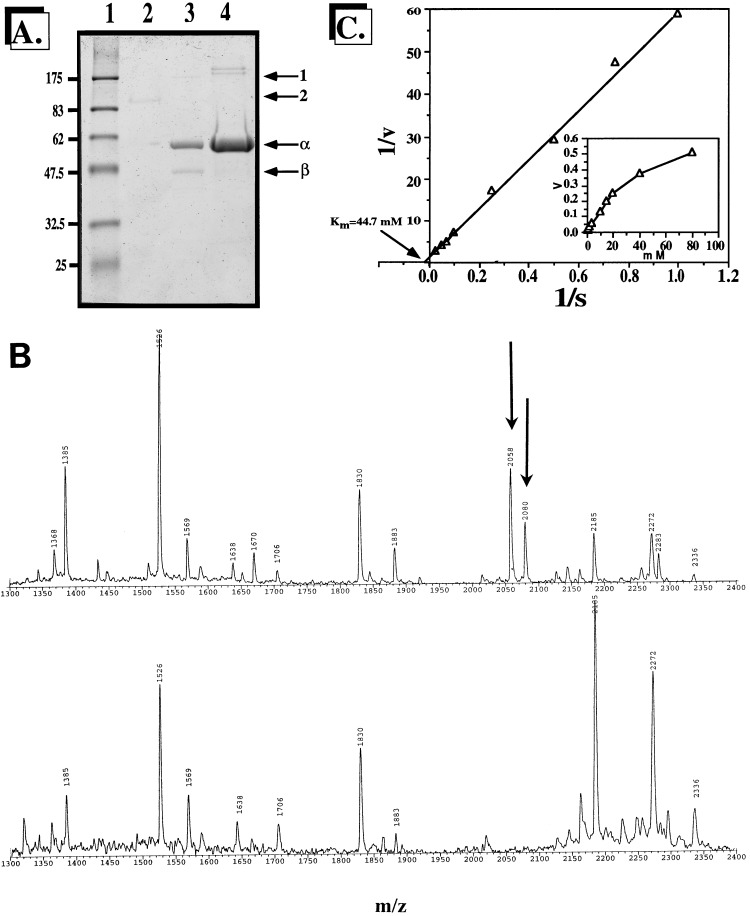
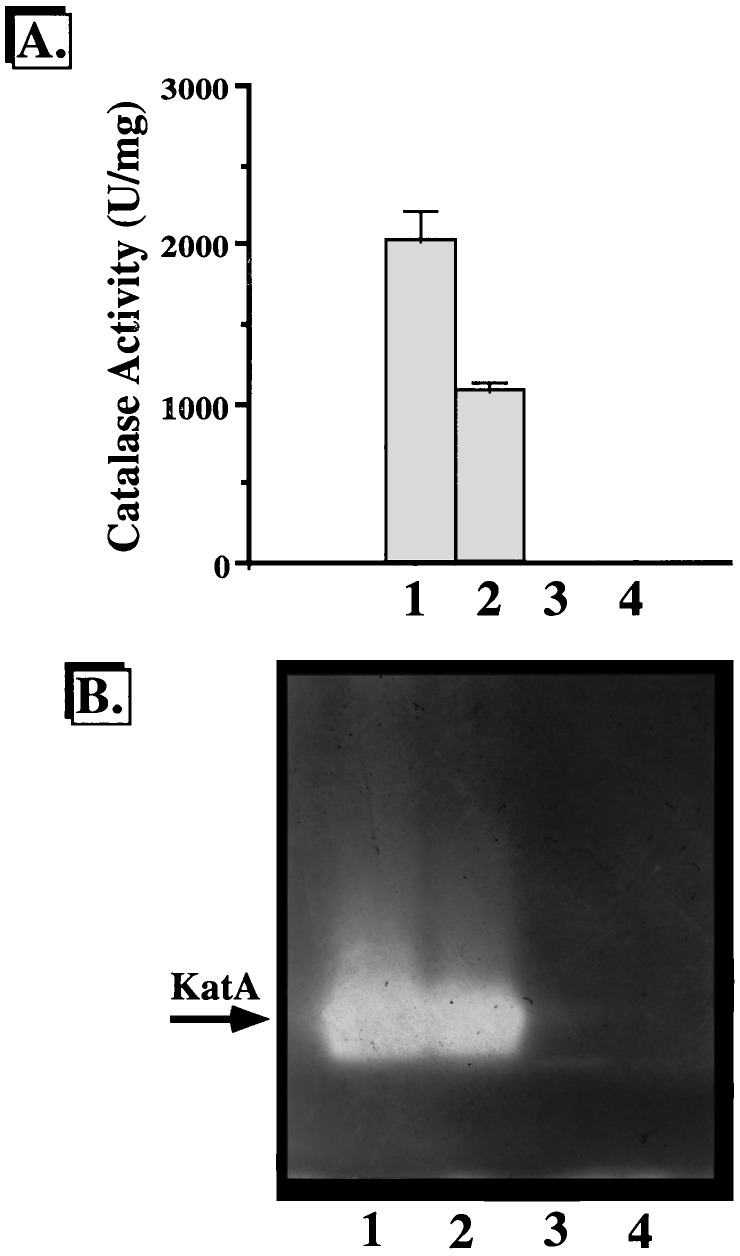
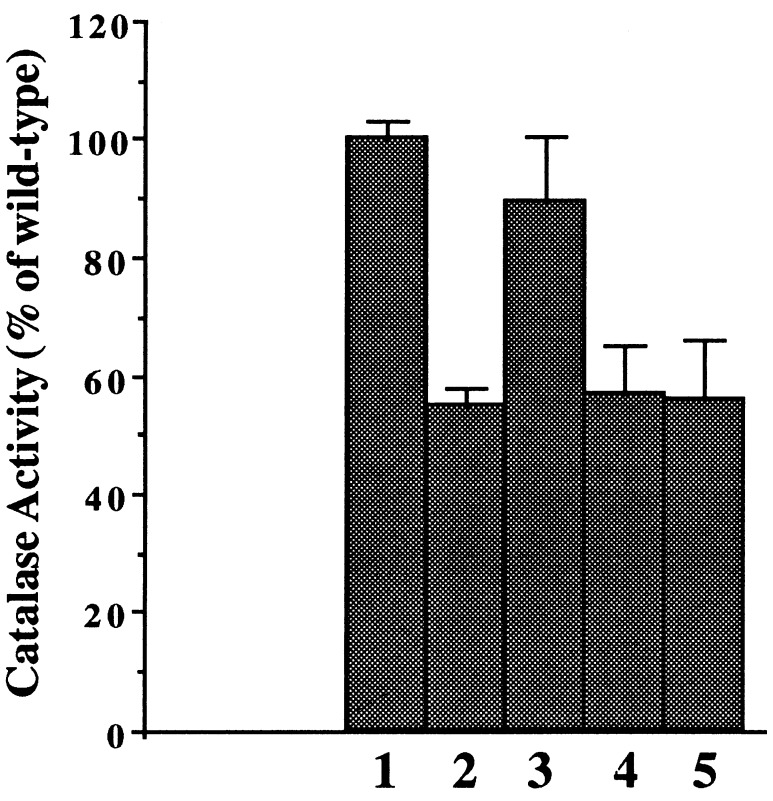
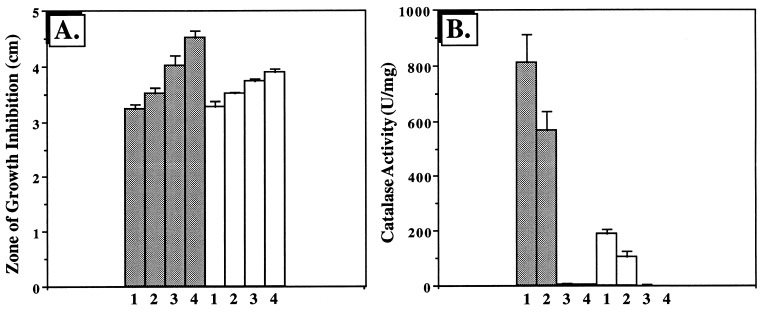
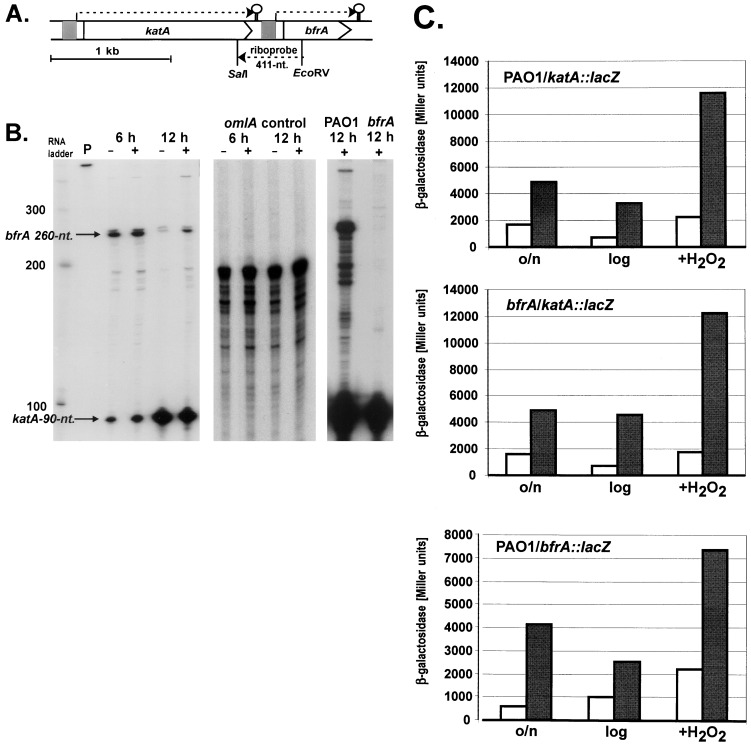
We have cloned a 3.6-kb genomic DNA fragment from Pseudomonas aeruginosa harboring the rpoA, rplQ, katA, and bfrA genes. These loci are predicted to encode, respectively, (i) the alpha subunit of RNA polymerase; (ii) the L17 ribosomal protein; (iii) the major catalase, KatA; and (iv) one of two iron storage proteins called bacterioferritin A (BfrA; cytochrome b1 or b557). Our goal was to determine the contributions of KatA and BfrA to the resistance of P. aeruginosa to hydrogen peroxide (H2O2). When provided on a multicopy plasmid, the P. aeruginosa katA gene complemented a catalase-deficient strain of Escherichia coli. The katA gene was found to contain two translational start codons encoding a heteromultimer of approximately 160 to 170 kDa and having an apparent Km for H2O2 of 44.7 mM. Isogenic katA and bfrA mutants were hypersusceptible to H2O2, while a katA bfrA double mutant demonstrated the greatest sensitivity. The katA and katA bfrA mutants possessed no detectable catalase activity. Interestingly, a bfrA mutant expressed only approximately 47% the KatA activity of wild-type organisms, despite possessing wild-type katA transcription and translation. Plasmids harboring bfrA genes encoding BfrA altered at critical amino acids essential for ferroxidase activity could not restore wild-type catalase activity in the bfrA mutant. RNase protection assays revealed that katA and bfrA are on different transcripts, the levels of which are increased by both iron and H2O2. Mass spectrometry analysis of whole cells revealed no significant difference in total cellular iron levels in the bfrA, katA, and katA bfrA mutants relative to wild-type bacteria. Our results suggest that P. aeruginosa BfrA may be required as one source of iron for the heme prosthetic group of KatA and thus for protection against H2O2.
Figures

References
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
-
- Andrews S C, Smith J M, Yewdall S J, Guest J R, Harrison P M. Bacterioferritins and ferritins are distantly related in evolution: conservation of ferroxidase-centre residues. FEBS Lett. 1991;293:164–168. - PubMed
-
- Ausubel F M, Brent R, Kingston R E, Moore D D, Seidman J D, Struhl K. Current protocols in molecular biology. New York, N.Y: John Wiley & Sons, Inc.; 1993. pp. 5.3.2–5.3.8.
-
- Bando Y, Aki K. Superoxide-mediated release of iron from ferritin by some flavoenzymes. Biochem Biophys Res Commun. 1990;168:389–395. - PubMed
-
- Beers R F, Jr, Sizer I W. A spectrophotometric method for measuring the breakdown of hydrogen peroxide by catalase. J Biol Chem. 1952;195:133–140. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
