

Transcriptional organization and in vivo role of the Escherichia coli rsd gene, encoding the regulator of RNA polymerase sigma D
- PMID: 10368152
- PMCID: PMC93855
- DOI: 10.1128/JB.181.12.3768-3776.1999
Transcriptional organization and in vivo role of the Escherichia coli rsd gene, encoding the regulator of RNA polymerase sigma D
Abstract
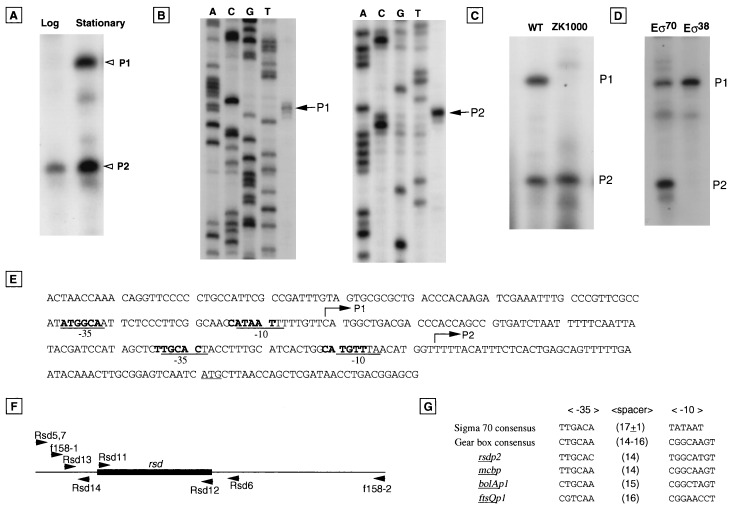
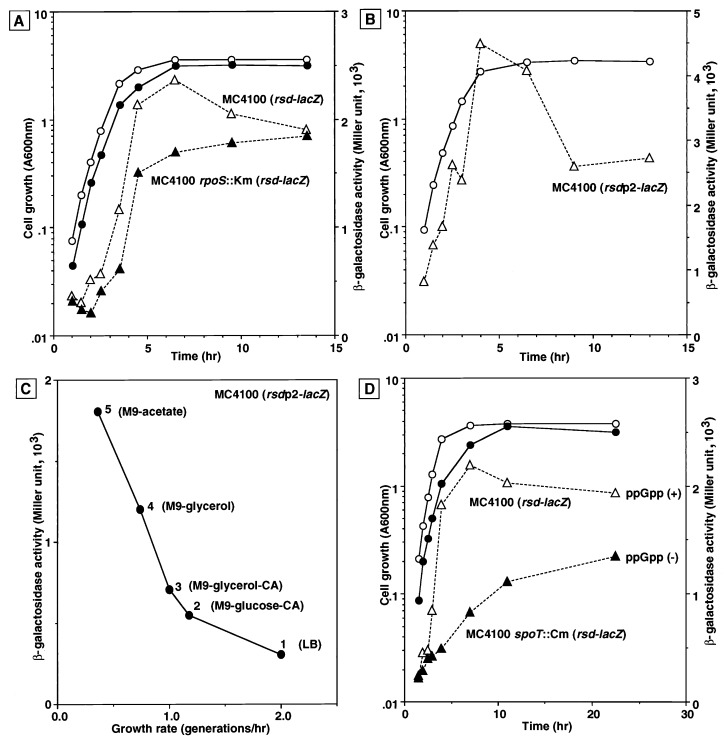
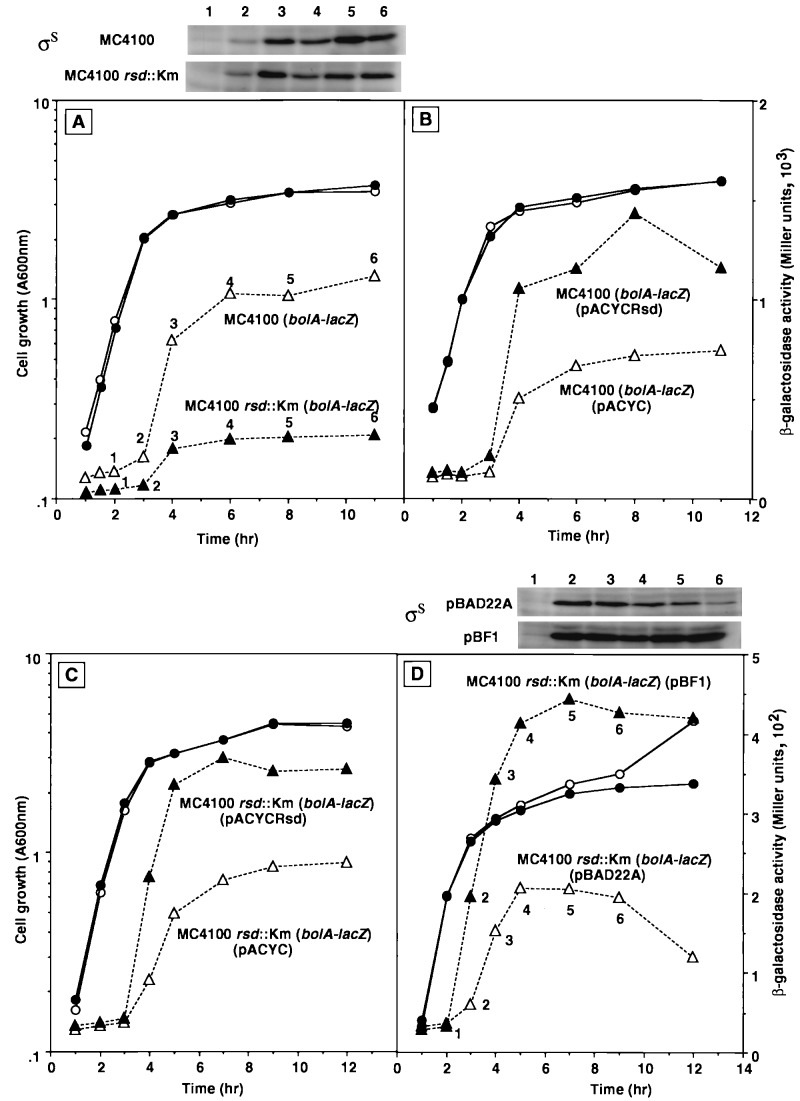
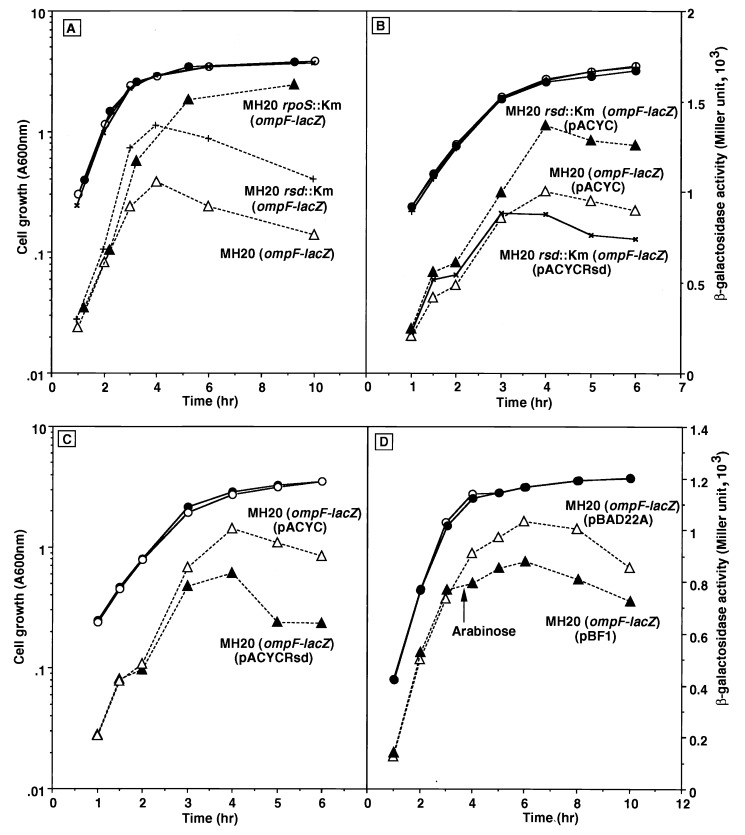
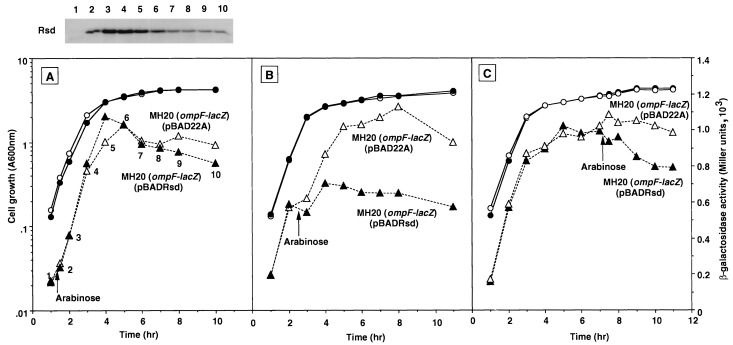
The regulator of sigma D (Rsd) was identified as an RNA polymerase sigma70-associated protein in stationary-phase Escherichia coli with the inhibitory activity of sigma70-dependent transcription in vitro (M. Jishage and A. Ishihama, Proc. Natl. Acad. Sci. USA 95:4953-4958, 1998). Primer extension analysis of rsd mRNA indicated the presence of two promoters, sigmaS-dependent P1 and sigma70-dependent P2 with the gearbox sequence. To get insight into the in vivo role of Rsd, the expression of a reporter gene fused to either the sigma70- or sigmaS-dependent promoter was analyzed in the absence of Rsd or the presence of overexpressed Rsd. In the rsd null mutant, the sigma70- and sigmaS-dependent gene expression was increased or decreased, respectively. On the other hand, the sigma70- or sigmaS-dependent transcription was reduced or enhanced, respectively, after overexpression of Rsd. The repression of the sigmaS-dependent transcription in the rsd mutant is overcome by increased production of the sigmaS subunit. Together these observations support the prediction that Rsd is involved in replacement of the RNA polymerase sigma subunit from sigma70 to sigmaS during the transition from exponential growth to the stationary phase.
Figures
Similar articles
-
A stationary phase protein in Escherichia coli with binding activity to the major sigma subunit of RNA polymerase.Proc Natl Acad Sci U S A. 1998 Apr 28;95(9):4953-8. doi: 10.1073/pnas.95.9.4953. Proc Natl Acad Sci U S A. 1998. PMID: 9560209 Free PMC article.
-
Negative regulation by RpoS: a case of sigma factor competition.Mol Microbiol. 1998 Aug;29(4):1039-51. doi: 10.1046/j.1365-2958.1998.00990.x. Mol Microbiol. 1998. PMID: 9767572
-
The Escherichia coli Ada protein can interact with two distinct determinants in the sigma70 subunit of RNA polymerase according to promoter architecture: identification of the target of Ada activation at the alkA promoter.J Bacteriol. 1999 Mar;181(5):1524-9. doi: 10.1128/JB.181.5.1524-1529.1999. J Bacteriol. 1999. PMID: 10049384 Free PMC article.
-
The molecular basis of selective promoter activation by the sigmaS subunit of RNA polymerase.Mol Microbiol. 2007 Mar;63(5):1296-306. doi: 10.1111/j.1365-2958.2007.05601.x. Mol Microbiol. 2007. PMID: 17302812 Review.
-
Recent insights into the general stress response regulatory network in Escherichia coli.J Mol Microbiol Biotechnol. 2002 May;4(3):341-6. J Mol Microbiol Biotechnol. 2002. PMID: 11931567 Review.
Cited by
-
Insight into the biology of Mycobacterium mucogenicum and Mycobacterium neoaurum clade members.Sci Rep. 2019 Dec 17;9(1):19259. doi: 10.1038/s41598-019-55464-5. Sci Rep. 2019. PMID: 31848383 Free PMC article.
-
PRODORIC: state-of-the-art database of prokaryotic gene regulation.Nucleic Acids Res. 2022 Jan 7;50(D1):D295-D302. doi: 10.1093/nar/gkab1110. Nucleic Acids Res. 2022. PMID: 34850133 Free PMC article.
-
Multiprobe RNase protection assay analysis of mRNA levels for the Escherichia coli oxidative DNA glycosylase genes under conditions of oxidative stress.J Bacteriol. 2000 Oct;182(19):5416-24. doi: 10.1128/JB.182.19.5416-5424.2000. J Bacteriol. 2000. PMID: 10986244 Free PMC article.
-
Binding of sigma(A) and sigma(B) to core RNA polymerase after environmental stress in Bacillus subtilis.J Bacteriol. 2003 Jan;185(1):35-40. doi: 10.1128/JB.185.1.35-40.2003. J Bacteriol. 2003. PMID: 12486038 Free PMC article.
-
Coordinated Hibernation of Transcriptional and Translational Apparatus during Growth Transition of Escherichia coli to Stationary Phase.mSystems. 2018 Sep 11;3(5):e00057-18. doi: 10.1128/mSystems.00057-18. eCollection 2018 Sep-Oct. mSystems. 2018. PMID: 30225374 Free PMC article.
References
-
- Aiba H, Adhya S, de Crombrugghe B. Evidence for two functional gal promoters in intact Escherichia colicells. J Biol Chem. 1981;256:11905–11910. - PubMed
-
- Angerer A, Enz S, Ochs M, Braun V. Transcriptional regulation of ferric citrate transport in Escherichia coli K-12. FecI belongs to a new subfamily of ς70-type factors that respond to extracytoplasmic stimuli. Mol Microbiol. 1995;18:163–174. - PubMed
-
- Ballesteros M, Kusano S, Ishihama A, Vicente M. The ftsQ1p gearbox promoter of Escherichia coli is a major sigma S-dependent promoter in the ddlB-ftsAregion. Mol Microbiol. 1998;30:419–430. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
