

Transcriptional regulation in the hyperthermophilic archaeon Pyrococcus furiosus: coordinated expression of divergently oriented genes in response to beta-linked glucose polymers
- PMID: 10368153
- PMCID: PMC93856
- DOI: 10.1128/JB.181.12.3777-3783.1999
Transcriptional regulation in the hyperthermophilic archaeon Pyrococcus furiosus: coordinated expression of divergently oriented genes in response to beta-linked glucose polymers
Abstract
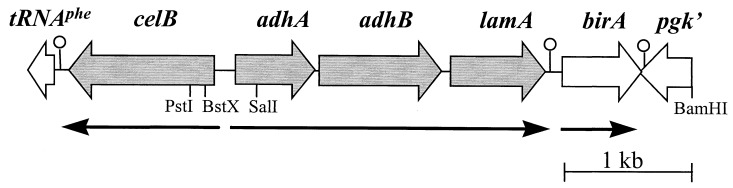
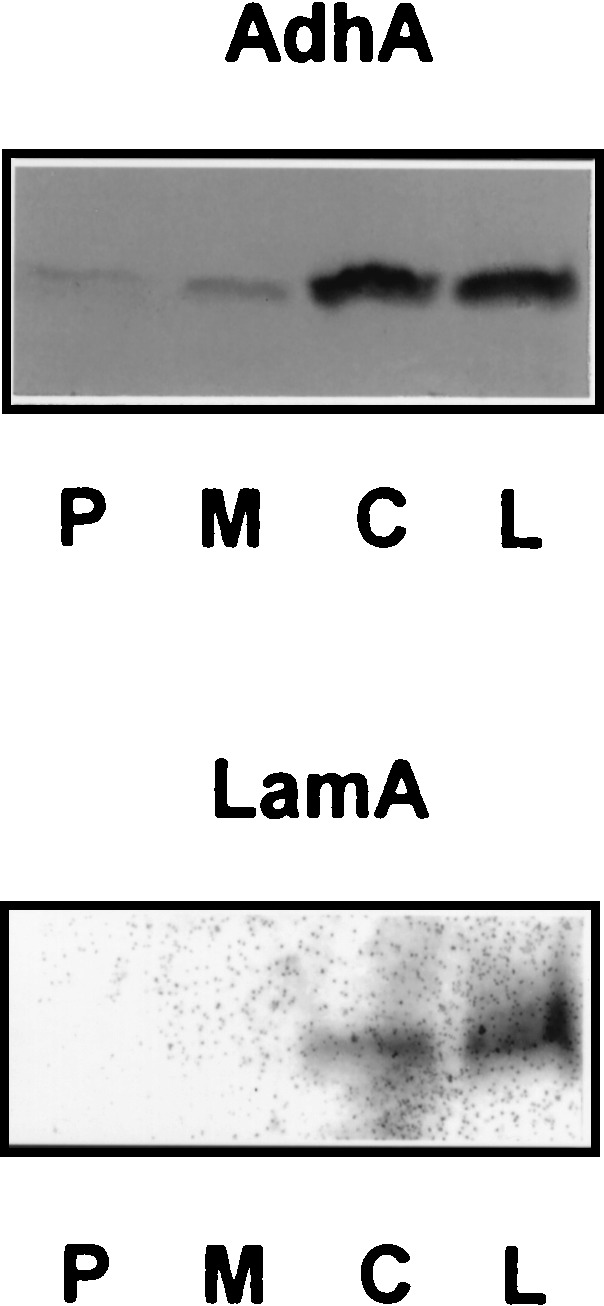
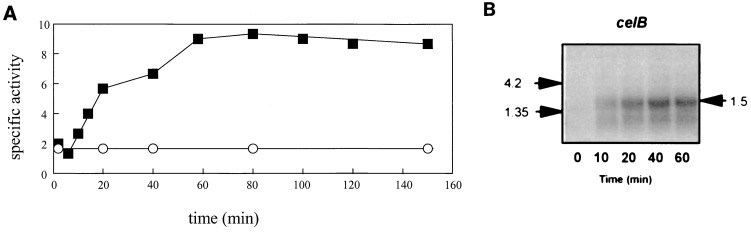
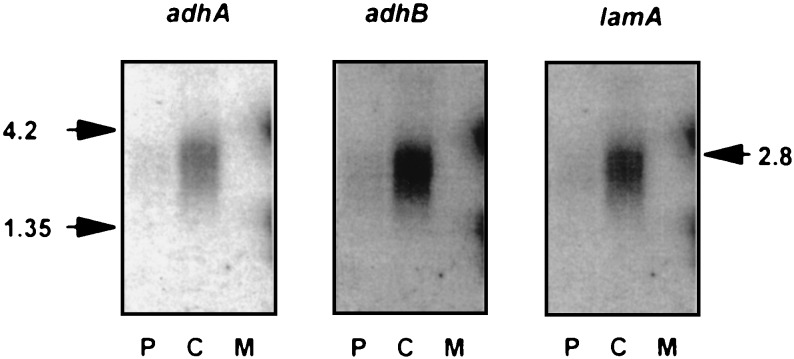
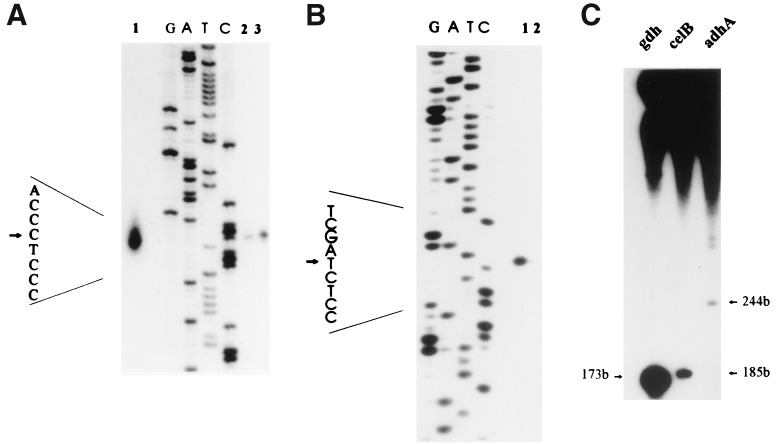
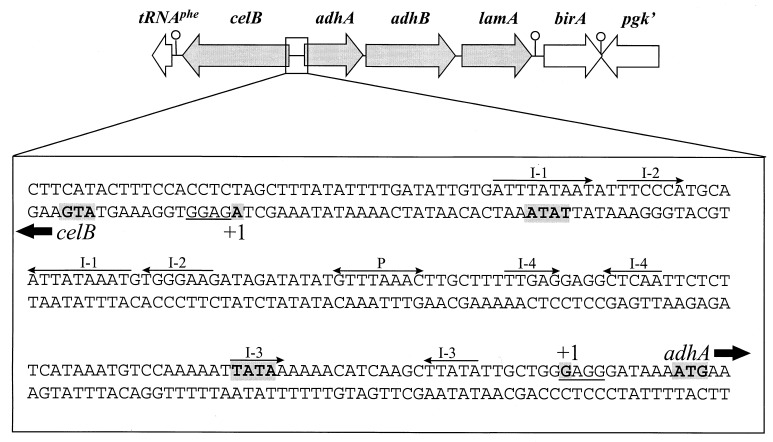
The genetic organization, expression, and regulation of the celB locus of the hyperthermophilic archaeon Pyrococcus furiosus were analyzed. This locus includes the celB gene, which codes for an intracellular beta-glucosidase, and a divergently orientated gene cluster, adhA-adhB-lamA, which codes for two alcohol dehydrogenases and an extracellular beta-1,3-endoglucanase that is transcribed as a polycistronic messenger (the lamA operon). During growth of P. furiosus on either the beta-1,4-linked glucose dimer cellobiose or the beta-1,3-linked glucose polymer laminarin, the activities of both beta-glucosidase and endoglucanase were increased at least fivefold compared with levels during growth on maltose or pyruvate. Northern blot analysis revealed an enhanced transcription of both the celB gene and the lamA operon in the presence of these glucose-containing substrates. The in vivo and in vitro transcription initiation sites of both the celB gene and the lamA operon were identified 25 nucleotides downstream of conserved TATA box motifs. A number of repeating sequences have been recognized in the celB-adhA intergenic region, some of which might be part of a transcriptional regulator-binding site.
Figures
References
-
- Baumann P, Qureshi S A, Jackson S P. Transcription: new insights from studies on Archaea. Trends Genet. 1995;11:279–283. - PubMed
-
- Bell S D, Jackson S P. Transcription and translation in Archaea: a mosaic of eukaryal and bacterial features. Trends Microbiol. 1998;6:222–228. - PubMed
-
- Brinkman, A. B., I. Dahlke, J. E. Tuininga, T. Lammers, V. Dumay, E. de Heus, J. H. G. Lebbink, M. Thomm, W. M. de Vos, and J. van der Oost. Submitted for publication. - PubMed
-
- Brown J W, Daniels C J, Reeve J N. Gene structure, organization, and expression in archaebacteria. Crit Rev Microbiol. 1989;16:287–338. - PubMed
-
- Bult C J, White O, Olsen G J, Zhou L X, Fleischmann R D, Sutton G G, Blake J A, FitzGerald L M, Clayton R A, Gocayne J D, Kerlavage A R, Dougherty B A, Tomb J F, Adams M D, Reich C I, Overbeek R, Kirkness E F, Weinstock K G, Merrick J M, Glodek A, Scott J L, Geoghagen N S M, Weidman J F, Fuhrmann J L, Nguyen D, Utterback T R, Kelley J M, Peterson J D, Sadow P W, Hanna M C, Cotton M D, Roberts K M, Hurst M A, Kaine B P, Borodovsky M, Klenk H P, Fraser C M, Smith H O, Woese C R, Venter J C. Complete genome sequence of the methanogenic archaeon, Methanococcus jannaschii. Science. 1996;273:1058–1073. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
