

Biochemical and molecular characterization of the Bacillus subtilis acetoin catabolic pathway
- PMID: 10368162
- PMCID: PMC93865
- DOI: 10.1128/JB.181.12.3837-3841.1999
Biochemical and molecular characterization of the Bacillus subtilis acetoin catabolic pathway
Abstract
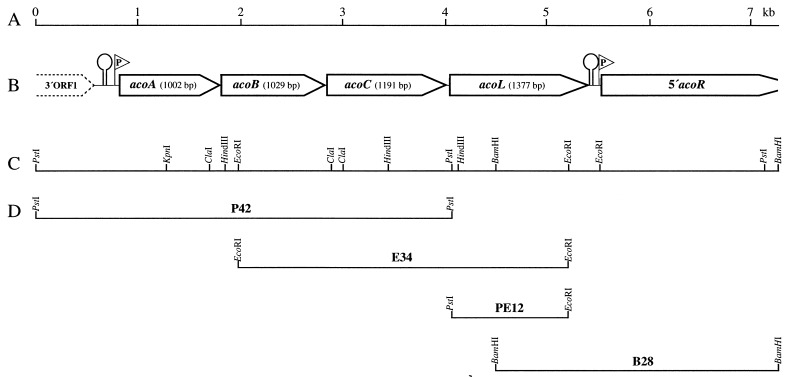
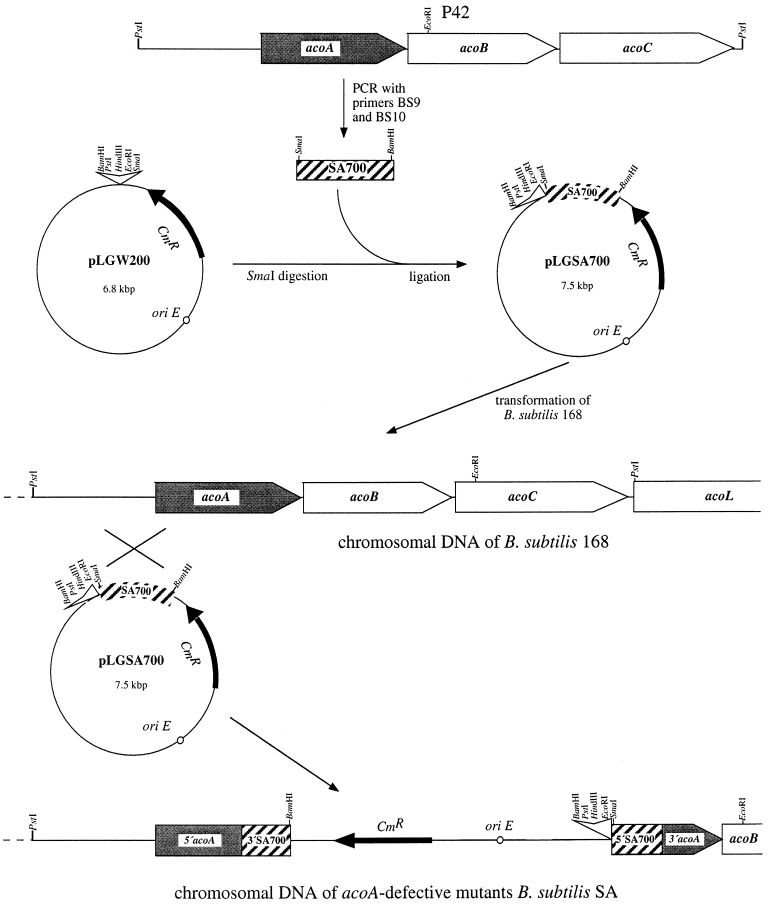
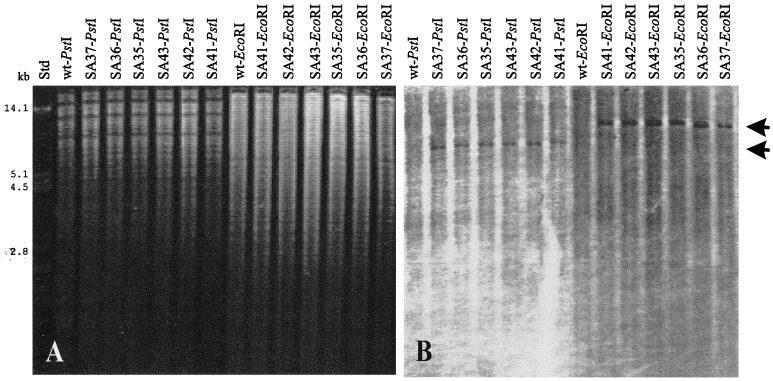
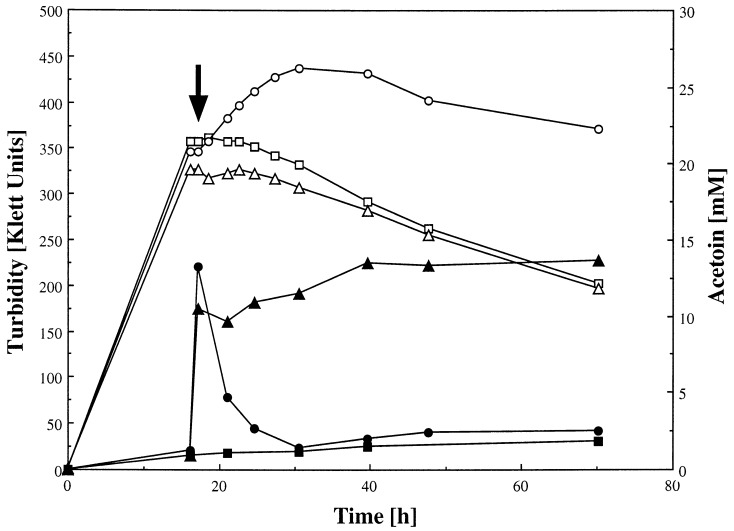
A recent study indicated that Bacillus subtilis catabolizes acetoin by enzymes encoded by the acu gene cluster (F. J. Grundy, D. A. Waters, T. Y. Takova, and T. M. Henkin, Mol. Microbiol. 10:259-271, 1993) that are completely different from those in the multicomponent acetoin dehydrogenase enzyme system (AoDH ES) encoded by aco gene clusters found before in all other bacteria capable of utilizing acetoin as the sole carbon source for growth. By hybridization with a DNA probe covering acoA and acoB of the AoDH ES from Clostridium magnum, genomic fragments from B. subtilis harboring acoA, acoB, acoC, acoL, and acoR homologous genes were identified, and some of them were functionally expressed in E. coli. Furthermore, acoA was inactivated in B. subtilis by disruptive mutagenesis; these mutants were impaired to express PPi-dependent AoDH E1 activity to remove acetoin from the medium and to grow with acetoin as the carbon source. Therefore, acetoin is catabolized in B. subtilis by the same mechanism as all other bacteria investigated so far, leaving the function of the previously described acu genes obscure.
Figures
References
-
- Bolhuis A, Sorokin A, Azevedo V, Ehrlich S D, Braun P G, de Jong A, Venema G, Bron S, van Dijl J M. Bacillus subtilis can modulate its capacity and specificity for protein secretion through temporally controlled expression of the sipSgene for signal peptidase. Mol Microbiol. 1996;22:605–618. - PubMed
-
- Bron S, Venema G. Ultraviolet inactivation and excision-repair in Bacillus subtilis. I. Construction and characterization of a transformable eightfold auxotrophic strain and two ultraviolet-sensitive derivatives. Mutat Res. 1972;15:1–10. - PubMed
-
- Browner M F, Taroni F, Sztul E, Rosenberg L E. Sequence analysis, biogenesis, and mitochondrial import of the α-subunit of rat propionyl-coA carboxylase. J Biol Chem. 1989;264:12680–12685. - PubMed
-
- Campbell A. Episomes. Adv Genet. 1962;11:101–145.
-
- Carothers D J, Pons G, Patel M S. Dihydrolipoamide dehydrogenase: functional similarities and divergent evolution of the pyridine nucleotide-disulfide oxidoreductases. Arch Biochem Biophys. 1989;268:409–425. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
