

A RAG1 and RAG2 tetramer complex is active in cleavage in V(D)J recombination
- PMID: 10373515
- PMCID: PMC84264
- DOI: 10.1128/MCB.19.7.4664
A RAG1 and RAG2 tetramer complex is active in cleavage in V(D)J recombination
Abstract
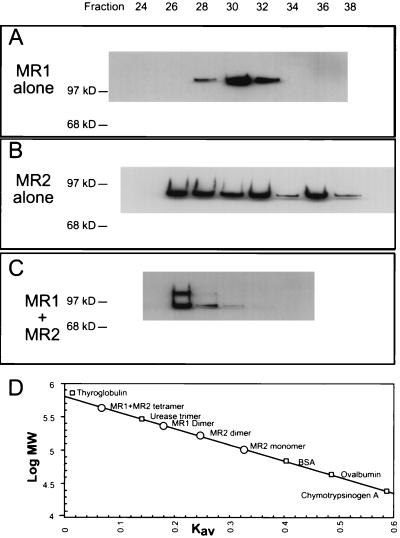
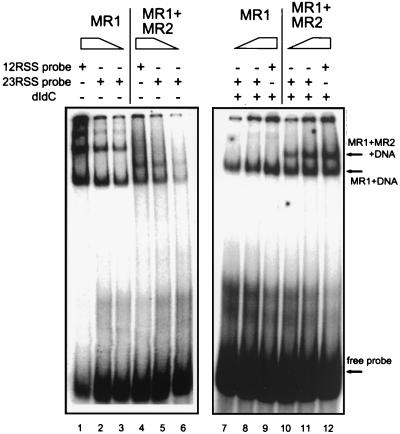
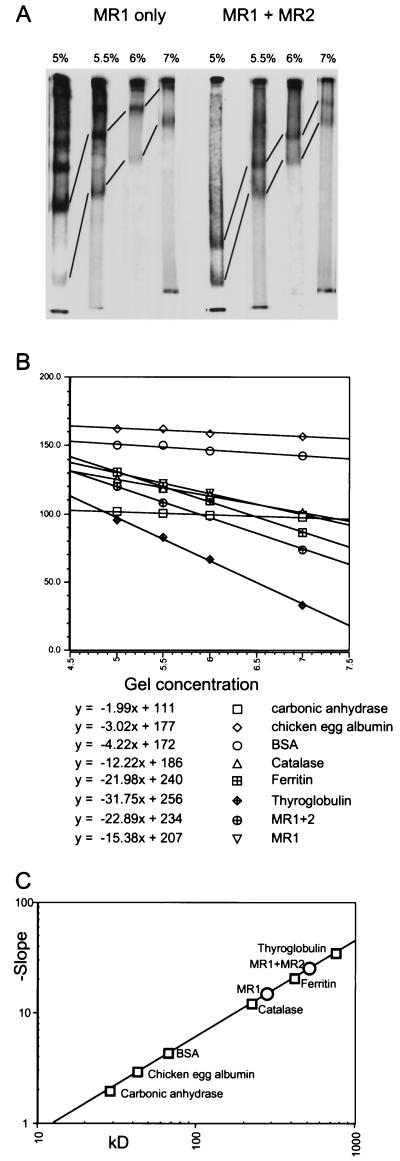
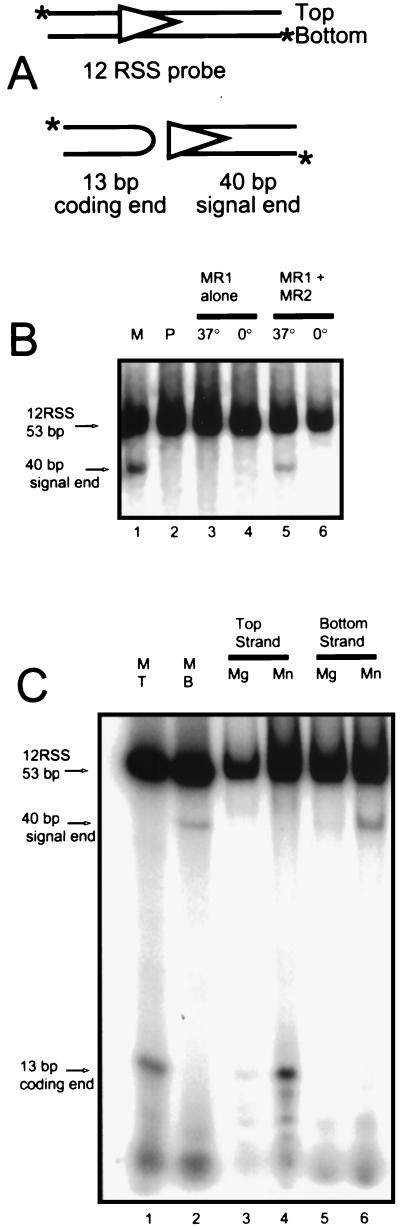
During V(D)J recombination two proteins, RAG1 and RAG2, assemble as a protein-DNA complex with the appropriate DNA targets containing recombination signal sequences (RSSs). The properties of this complex require a fairly elaborate set of protein-protein and protein-DNA contacts. Here we show that a purified derivative of RAG1, without DNA, exists predominantly as a homodimer. A RAG2 derivative alone has monomer, dimer, and larger forms. The coexpressed RAG1 and RAG2 proteins form a mixed tetramer in solution which contains two molecules of each protein. The same tetramer of RAG1 and RAG2 plus one DNA molecule is the form active in cleavage. Additionally, we show that both DNA products following cleavage can still be held together in a stable protein-DNA complex.
Figures
References
-
- Besmer E, Mansilla-Soto J, Cassard S, Sawchuk D J, Brown G, Sadofsky M, Lewis S M, Nussenzweig M C, Cortes P. Hairpin coding end opening is mediated by RAG1 and RAG2 proteins. Mol Cell. 1998;2:817–828. - PubMed
-
- Bullock B P, Habener J F. Phosphorylation of the cAMP response element binding protein CREB by cAMP-dependent protein kinase A and glycogen synthase kinase-3 alters DNA-binding affinity, conformation, and increases net charge. Biochemistry. 1998;37:3795–3809. - PubMed
-
- Difilippantonio M J, McMahan C J, Eastman Q M, Spanopoulou E, Schatz D G. RAG1 mediates signal sequence recognition and recruitment of RAG2 in V(D)J recombination. Cell. 1996;87:253–262. - PubMed
-
- Eastman Q M, Leu T M J, Schatz D G. Initiation of V(D)J recombination in-vitro obeying the 12/23-rule. Nature. 1996;380:85–88. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources