

Atm inactivation results in aberrant telomere clustering during meiotic prophase
- PMID: 10373558
- PMCID: PMC84352
- DOI: 10.1128/MCB.19.7.5096
Atm inactivation results in aberrant telomere clustering during meiotic prophase
Abstract
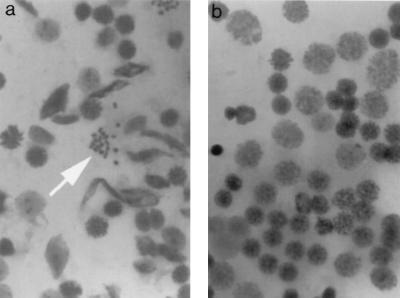
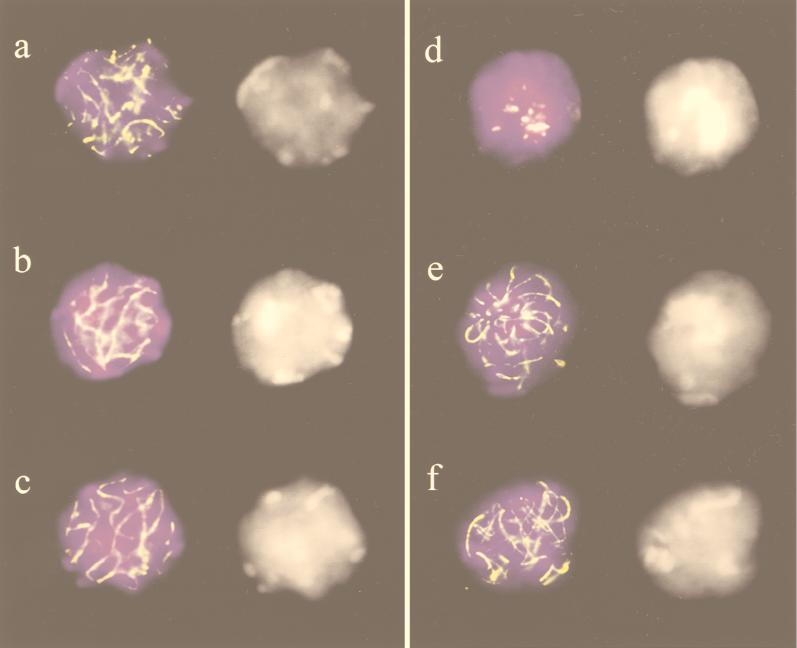
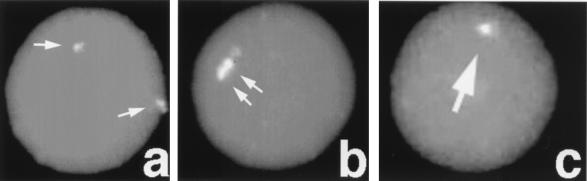
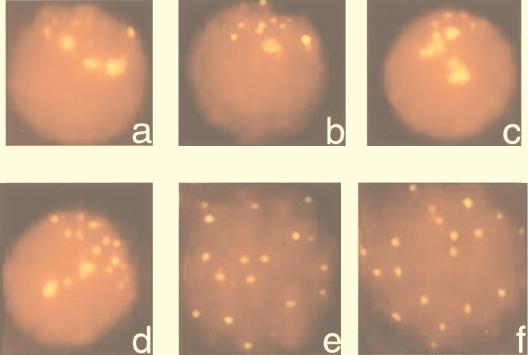
A-T (ataxia telangiectasia) individuals frequently display gonadal atrophy, and Atm-/- mice show spermatogenic failure due to arrest at prophase of meiosis I. Chromosomal movements take place during meiotic prophase, with telomeres congregating on the nuclear envelope to transiently form a cluster during the leptotene/zygotene transition (bouquet arrangement). Since the ATM protein has been implicated in telomere metabolism of somatic cells, we have set out to investigate the effects of Atm inactivation on meiotic telomere behavior. Fluorescent in situ hybridization and synaptonemal complex (SC) immunostaining of structurally preserved spermatocytes I revealed that telomere clustering occurs aberrantly in Atm-/- mice. Numerous spermatocytes of Atm-/- mice displayed locally accumulated telomeres with stretches of SC near the clustered chromosome ends. This contrasted with spermatogenesis of normal mice, where only a few leptotene/zygotene spermatocytes I with clustered telomeres were detected. Pachytene nuclei, which were much more abundant in normal mice, displayed telomeres scattered over the nuclear periphery. It appears that the timing and occurrence of chromosome polarization is altered in Atm-/- mice. When we examined telomere-nuclear matrix interactions in spermatocytes I, a significant difference was observed in the ratio of soluble versus matrix-associated telomeric DNA sequences between meiocytes of Atm-/- and control mice. We propose that the severe disruption of spermatogenesis during early prophase I in the absence of functional Atm may be partly due to altered interactions of telomeres with the nuclear matrix and distorted meiotic telomere clustering.
Figures



Similar articles
-
Meiotic telomere distribution and Sertoli cell nuclear architecture are altered in Atm- and Atm-p53-deficient mice.Mol Cell Biol. 2000 Oct;20(20):7773-83. doi: 10.1128/MCB.20.20.7773-7783.2000. Mol Cell Biol. 2000. PMID: 11003672 Free PMC article.
-
Influence of ATM function on interactions between telomeres and nuclear matrix.Radiat Res. 2000 Aug;154(2):133-9. doi: 10.1667/0033-7587(2000)154[0133:ioafoi]2.0.co;2. Radiat Res. 2000. PMID: 10931683
-
ATM and RPA in meiotic chromosome synapsis and recombination.Nat Genet. 1997 Dec;17(4):457-61. doi: 10.1038/ng1297-457. Nat Genet. 1997. PMID: 9398850
-
Analysis of telomere dynamics in mouse spermatogenesis.Methods Mol Biol. 2009;558:383-99. doi: 10.1007/978-1-60761-103-5_22. Methods Mol Biol. 2009. PMID: 19685336 Review.
-
Knockout mice provide novel insights into meiotic chromosome and telomere dynamics.Cytogenet Genome Res. 2003;103(3-4):235-44. doi: 10.1159/000076809. Cytogenet Genome Res. 2003. PMID: 15051944 Review.
Cited by
-
Targeted inhibition of ATR or CHEK1 reverses radioresistance in oral squamous cell carcinoma cells with distal chromosome arm 11q loss.Genes Chromosomes Cancer. 2014 Feb;53(2):129-43. doi: 10.1002/gcc.22125. Epub 2013 Nov 25. Genes Chromosomes Cancer. 2014. PMID: 24327542 Free PMC article.
-
Tetrahymena meiotic nuclear reorganization is induced by a checkpoint kinase-dependent response to DNA damage.Mol Biol Cell. 2009 May;20(9):2428-37. doi: 10.1091/mbc.e08-10-1058. Epub 2009 Mar 18. Mol Biol Cell. 2009. PMID: 19297526 Free PMC article.
-
ATM-dependent DNA damage response constrains cell growth and drives clonal hematopoiesis in telomere biology disorders.J Clin Invest. 2025 Apr 3;135(8):e181659. doi: 10.1172/JCI181659. eCollection 2025 Apr 15. J Clin Invest. 2025. PMID: 40179146 Free PMC article.
-
FBXO47 regulates telomere-inner nuclear envelope integration by stabilizing TRF2 during meiosis.Nucleic Acids Res. 2019 Dec 16;47(22):11755-11770. doi: 10.1093/nar/gkz992. Nucleic Acids Res. 2019. PMID: 31724724 Free PMC article.
-
Meiosis-specific cohesin mediates homolog recognition in mouse spermatocytes.Genes Dev. 2014 Mar 15;28(6):594-607. doi: 10.1101/gad.237313.113. Epub 2014 Mar 3. Genes Dev. 2014. PMID: 24589552 Free PMC article.
References
-
- Ashley T. Mammalian meiotic recombination: a reexamination. Hum Genet. 1994;94:587–593. - PubMed
-
- Barlow C, Hirotsune S, Paylor R, Liyanage M, Eckhaus M, Collins F, Shiloh Y, Crawley J N, Ried T, Tagel D, Wynshaw-Boris A. Atm-deficient mice: a paradigm of ataxia-telangiectasia. Cell. 1996;86:159–171. - PubMed
-
- Barlow C, Liyanage M, Moens P B, Deng C-X, Ried T, Wynshaw-Boris A. Partial rescue of the prophase 1 defects of Atm-deficient mice by p53 and p21 null alleles. Nat Genet. 1997;17:462–466. - PubMed
-
- Barlow C, Liyanage M, Moens P B, Tarsounas M, Nagashima K, Brown K, Rottinghaus S P, Jackson S P, Tagle D, Ried T, Wynshaw-Boris A. Atm deficiency results in severe meiotic disruption as early as leptonema of prophase. Development. 1998;125:4007–4017. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Research Materials
Miscellaneous