

In vitro recordings of afferent fibres with receptive fields in the serosa, muscle and mucosa of rat colon
- PMID: 10373708
- PMCID: PMC2269405
- DOI: 10.1111/j.1469-7793.1999.0271r.x
In vitro recordings of afferent fibres with receptive fields in the serosa, muscle and mucosa of rat colon
Abstract
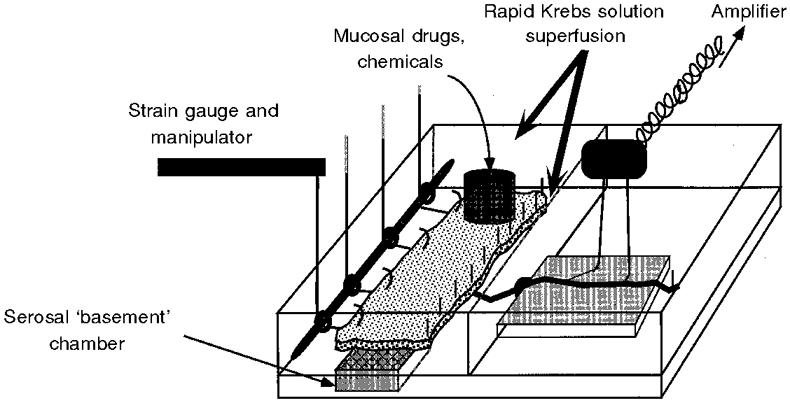
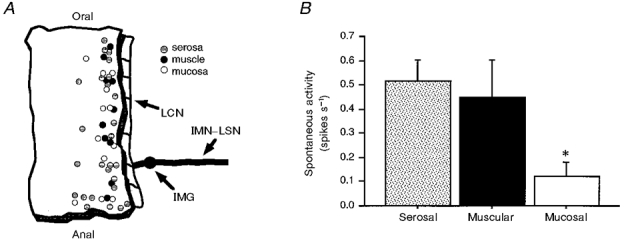
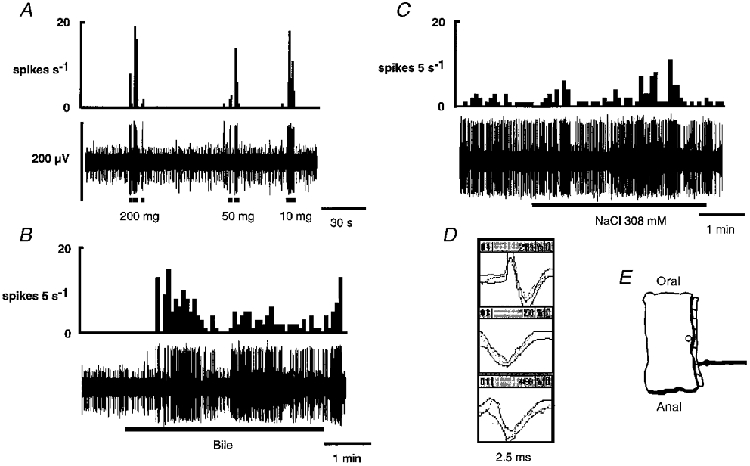
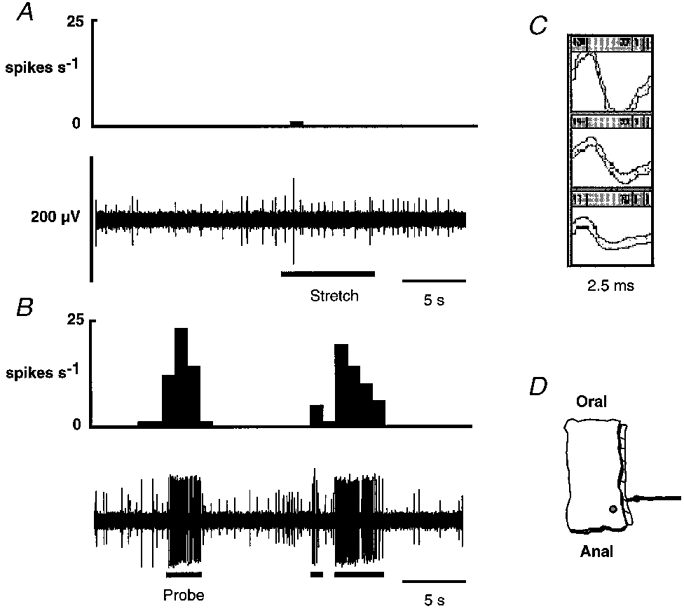
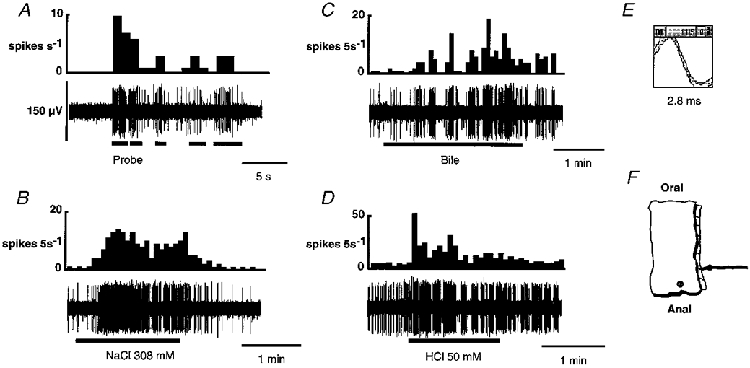
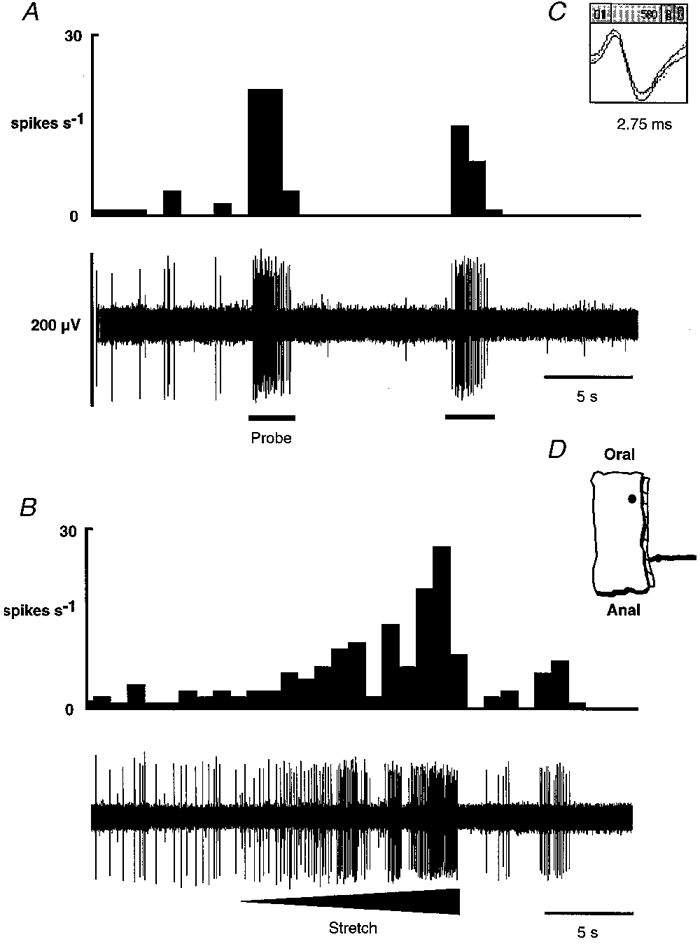
1. Colonic afferent fibres were recorded using a novel in vitro preparation. Fibres with endings in the colonic mucosa are described, along with those in muscle and serosa, and their responses to a range of mechanical and chemical luminal stimuli. 2. Mechanical stimuli were applied to the tissue, which included stretch, blunt probing of the mucosa and stroking of the mucosa with von Frey hairs (10-1000 mg). Chemical stimuli were applied into a ring that was placed over the mechanoreceptive field of the fibre; these were distilled water, 154 and 308 mM NaCl, 100 microM capsaicin, 50 mM HCl, and undiluted and 50% ferret bile. 3. Recordings were made from 52 fibres, 12 of which showed characteristics of having endings in the mucosa. Mucosal afferents were sensitive to a 10 mg von Frey hair and were generally chemosensitive to >= 1 chemical stimulus. 4. Ten fibres showed characteristics of having receptive fields in the muscular layer. These fibres responded readily to circumferential stretch, as well as to blunt probing. 5. Twenty-seven fibres showed characteristics of having endings in the serosal layer. They adapted rapidly to circumferential stretch and responded to blunt probing of the serosa. Fifteen of 19 serosal fibres tested also responded to luminal chemicals. 6. Three fibres were unresponsive to all mechanical stimuli but were recruited by chemical stimuli. 7. This is the first characterization of colonic afferent fibres using an in vitro method and the first documentation of afferent fibres with their endings in the mucosa of the colon. These fibres are likely to be important in aspects of colonic sensation and reflex control.
Figures
Similar articles
-
Vagal and spinal mechanosensors in the rat stomach and colon have multiple receptive fields.Am J Physiol Regul Integr Comp Physiol. 2001 May;280(5):R1371-81. doi: 10.1152/ajpregu.2001.280.5.R1371. Am J Physiol Regul Integr Comp Physiol. 2001. PMID: 11294756
-
An in vitro study of the properties of vagal afferent fibres innervating the ferret oesophagus and stomach.J Physiol. 1998 Nov 1;512 ( Pt 3)(Pt 3):907-16. doi: 10.1111/j.1469-7793.1998.907bd.x. J Physiol. 1998. PMID: 9769431 Free PMC article.
-
Properties of the major classes of mechanoreceptors in the guinea pig bladder.J Physiol. 2007 Nov 15;585(Pt 1):147-63. doi: 10.1113/jphysiol.2007.140244. Epub 2007 Oct 4. J Physiol. 2007. PMID: 17916614 Free PMC article.
-
Speculations on the structure/function relationship for vagal and splanchnic afferent endings supplying the gastrointestinal tract.J Auton Nerv Syst. 1988 Apr;22(3):175-80. doi: 10.1016/0165-1838(88)90104-x. J Auton Nerv Syst. 1988. PMID: 3047201 Review.
-
Airway nerves: in vitro electrophysiology.Curr Opin Pharmacol. 2002 Jun;2(3):278-9. doi: 10.1016/s1471-4892(02)00161-3. Curr Opin Pharmacol. 2002. PMID: 12020469 Review.
Cited by
-
P2X purinoceptor-induced sensitization of ferret vagal mechanoreceptors in oesophageal inflammation.J Physiol. 2000 Mar 1;523 Pt 2(Pt 2):403-11. doi: 10.1111/j.1469-7793.2000.00403.x. J Physiol. 2000. PMID: 10699084 Free PMC article.
-
Neuroanatomy of lower gastrointestinal pain disorders.World J Gastroenterol. 2014 Jan 28;20(4):1005-20. doi: 10.3748/wjg.v20.i4.1005. World J Gastroenterol. 2014. PMID: 24574773 Free PMC article. Review.
-
Development, plasticity and modulation of visceral afferents.Brain Res Rev. 2009 Apr;60(1):171-86. doi: 10.1016/j.brainresrev.2008.12.004. Epub 2008 Dec 25. Brain Res Rev. 2009. PMID: 19150371 Free PMC article. Review.
-
Differential chemosensory function and receptor expression of splanchnic and pelvic colonic afferents in mice.J Physiol. 2005 Aug 15;567(Pt 1):267-81. doi: 10.1113/jphysiol.2005.089714. Epub 2005 Jun 9. J Physiol. 2005. PMID: 15946967 Free PMC article.
-
Adrenergic stimulation mediates visceral hypersensitivity to colorectal distension following heterotypic chronic stress.Gastroenterology. 2010 Jan;138(1):294-304.e3. doi: 10.1053/j.gastro.2009.09.054. Epub 2009 Oct 1. Gastroenterology. 2010. PMID: 19800336 Free PMC article.
References
-
- Altschuler SM, Escardo J, Lynn RB, Miselis RR. The central organization of the vagus nerve innervating the colon of the rat. Gastroenterology. 1993;104:502–509. - PubMed
-
- Anthony TL, Kreulen DL. Volume-sensitive synaptic input to neurons in guinea pig inferior mesenteric ganglion. American Journal of Physiology. 1990;259:G490–497. - PubMed
-
- Baron R, Jänig W. Afferent and sympathetic neurons projecting into lumbar visceral nerves of the male rat. Journal of Comparative Neurology. 1991;314:429–436. - PubMed
-
- Baron R, Jänig W, Kollmann W. Sympathetic and afferent somata projecting in hindlimb nerves and the anatomical organization of the lumbar sympathetic nervous system of the rat. Journal of Comparative Neurology. 1988;275:460–468. - PubMed
-
- Berthoud HR, Carlson NR, Powley TL. Topography of efferent vagal innervation of the rat gastrointestinal tract. American Journal of Physiology. 1991;260:R200–207. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources