

Inhibition of TGF-beta 3 restores the invasive capability of extravillous trophoblasts in preeclamptic pregnancies
- PMID: 10377170
- PMCID: PMC408387
- DOI: 10.1172/JCI6380
Inhibition of TGF-beta 3 restores the invasive capability of extravillous trophoblasts in preeclamptic pregnancies
Abstract
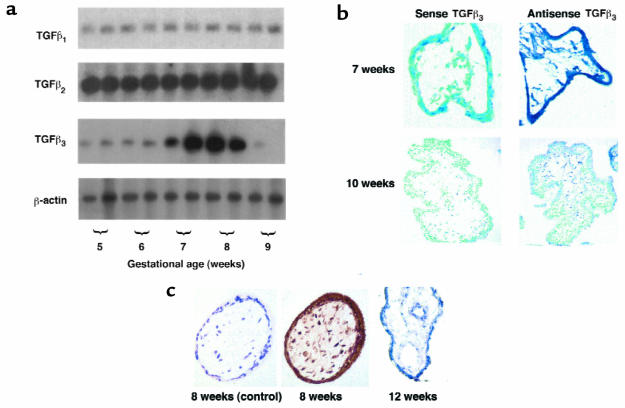
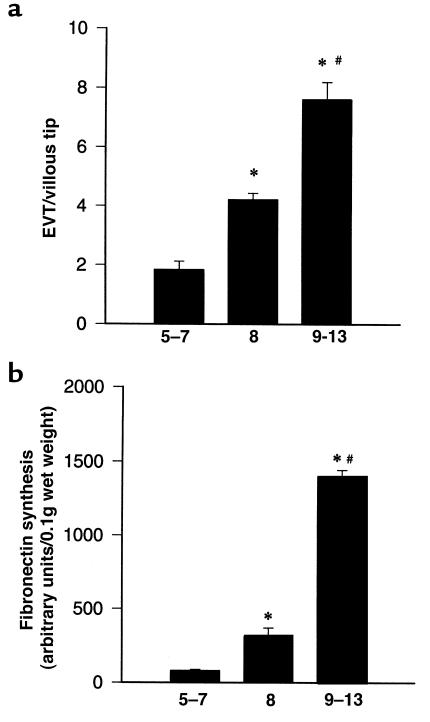
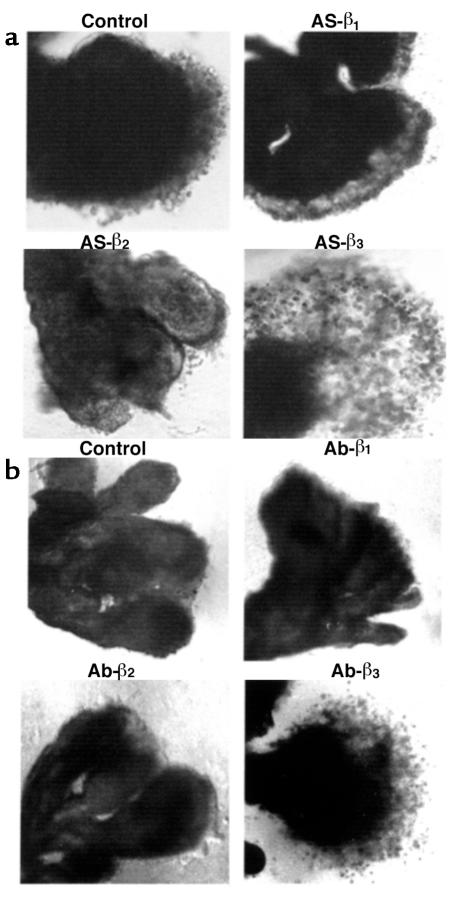
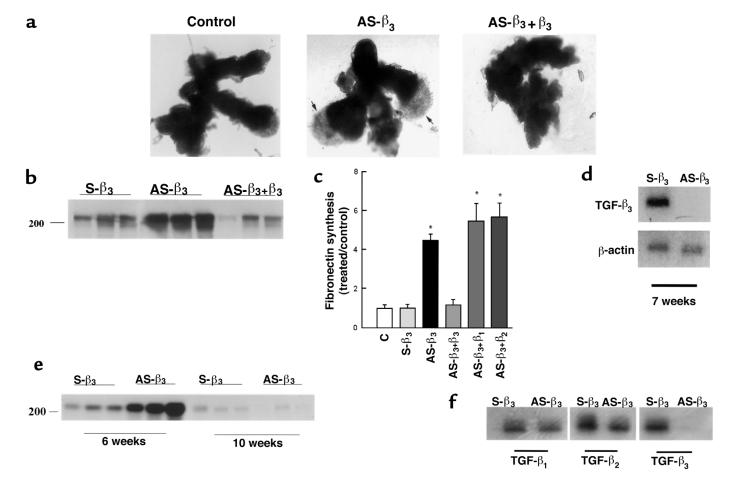
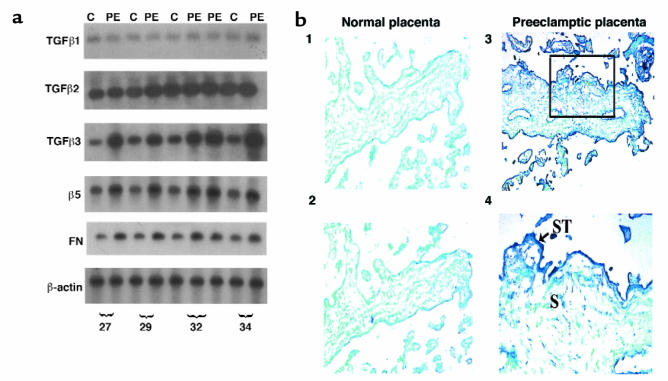
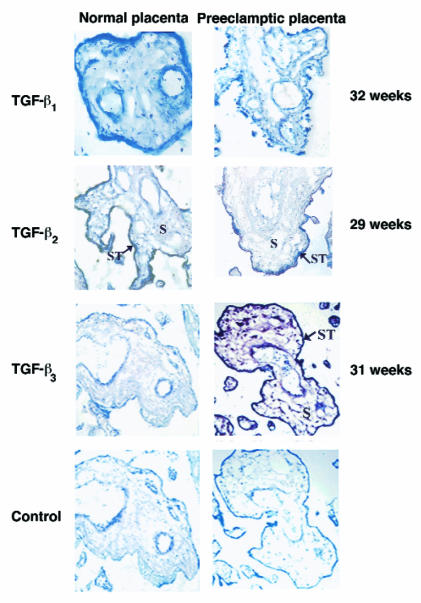
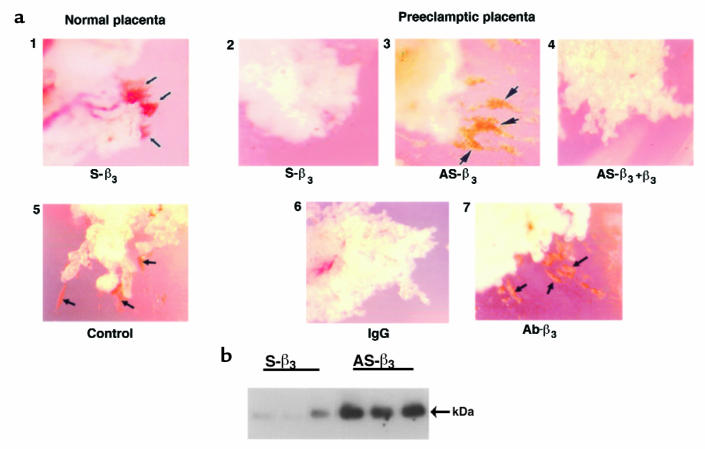
Preeclampsia, the major cause of maternal morbidity and mortality in developed countries, is associated with abnormalities of placenta function due to shallow invasion of the maternal decidua by trophoblasts. Data suggest that TGF-beta may play a role in inhibiting trophoblast outgrowth or invasion, or both. We report that placental TGF-beta 3 expression is high in early pregnancy but falls at around 9 weeks' gestation. This pattern is inversely correlated with trophoblast outgrowth and fibronectin synthesis, markers of early trophoblast differentiation toward an invasive phenotype. We demonstrate that TGF-beta 3 is overexpressed in preeclamptic placentae. In contrast to control placentae, explants from preeclamptic pregnancies fail to exhibit spontaneous invasion in vitro. Significantly, antisense-induced inhibition of TGF-beta 3 expression, and inhibition of TGF-beta 3 activity with antibodies, induces the formation of columns of trophoblast cells, which migrate out of the explant into the underlying Matrigel. To our knowledge, this is the first demonstration that the hypoinvasive placental phenotype characteristic of preeclampsia can be essentially normalized in vitro by biochemical manipulation. We speculate that a failure to downregulate expression of TGF-beta 3 at around 9 weeks' gestation results in shallow trophoblast invasion and predisposes the pregnancy to preeclampsia.
Figures

References
-
- Cross JC, Werb Z, Fisher SJ. Implantation and the placenta: key pieces of the development puzzle. Science. 1994;266:1508–1518. - PubMed
-
- Strickland S, Richards WG. Invasion of trophoblasts. Cell. 1992;71:355–357. - PubMed
-
- Aplin JD. Implantation, trophoblast differentiation and haemochorial placentation: mechanistic evidence in vivo and in vitro. J Cell Sci. 1991;99:681–692. - PubMed
-
- Redman CWG. Current topic: pre-eclampsia and the placenta. Placenta. 1991;12:301–308. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
