

Murine gammaherpesvirus M2 gene is latency-associated and its protein a target for CD8(+) T lymphocytes
- PMID: 10377445
- PMCID: PMC22116
- DOI: 10.1073/pnas.96.13.7508
Murine gammaherpesvirus M2 gene is latency-associated and its protein a target for CD8(+) T lymphocytes
Abstract
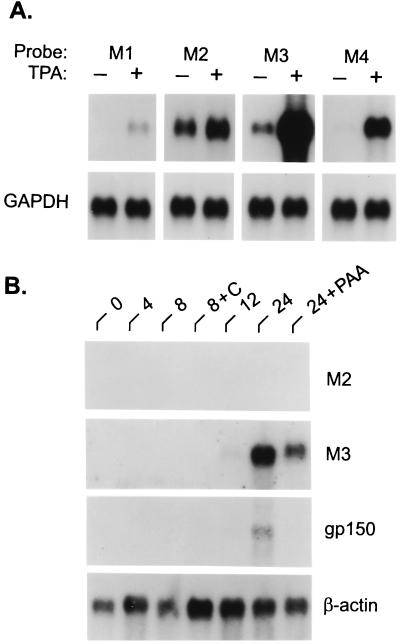
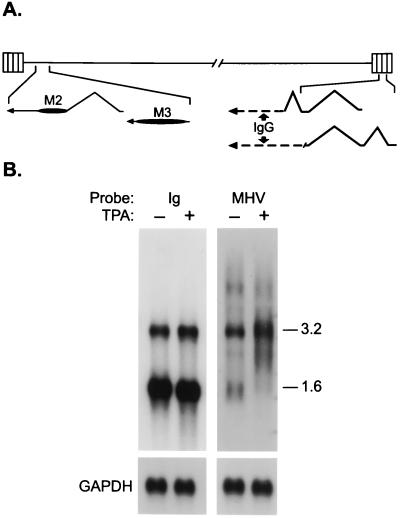
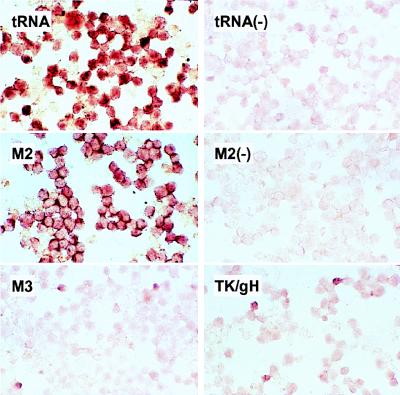
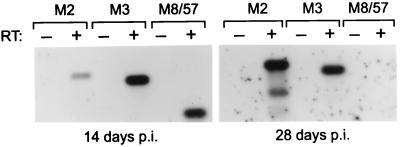
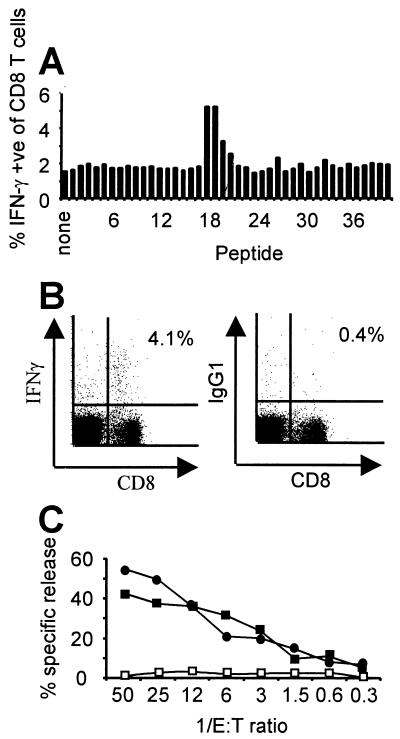
Murine gammaherpesvirus 68 (MHV-68) infection of mice is a potential model with which to address fundamental aspects of the pathobiology and host control of gammaherpesvirus latency. Control of MHV-68 infection, like that of Epstein-Barr virus, is strongly dependent on the cellular immune system. However, the molecular biology of MHV-68 latency is largely undefined. A screen of the MHV-68 genome for potential latency-associated mRNAs revealed that the region encompassing and flanking the genomic terminal repeats is transcriptionally active in the latently infected murine B-cell tumor line S11. Transcription of one MHV-68 gene, that encoding the hypothetical M2 protein, was detected in virtually all latently infected S11 cells and in splenocytes of latently infected mice, but not in lytically infected fibroblasts. Furthermore, an epitope was identified in the predicted M2 protein that is recognized by CD8(+) T cells from infected mice and a cytotoxic T lymphocyte line that recognizes this epitope killed S11 cells, indicating that the M2 protein is expressed during latent infection and is a target for the host cytotoxic T lymphocyte response. This work therefore provides essential information for modeling MHV-68 latency and strategies of immunotherapy against gammaherpesvirus-related diseases in a highly tractable animal model.
Figures
References
-
- Rickinson A B, Kieff E. In: Fields Virology. Fields B N, Knipe D M, Howley P M, editors. New York: Lippincott; 1996. pp. 2397–2446.
-
- Ganem D. Cell. 1997;91:157–160. - PubMed
-
- Boshoff C, Weiss R A. In: Advances in Cancer Research. Vande Woude G F, Klein G, editors. London: Academic; 1998. pp. 57–86. - PubMed
-
- Kieff E. In: Fields Virology. Fields B N, Knipe D M, Howley P M, editors. New York: Lippincott; 1996. pp. 2343–2396.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
