

BiP and immunoglobulin light chain cooperate to control the folding of heavy chain and ensure the fidelity of immunoglobulin assembly
- PMID: 10397760
- PMCID: PMC25436
- DOI: 10.1091/mbc.10.7.2209
BiP and immunoglobulin light chain cooperate to control the folding of heavy chain and ensure the fidelity of immunoglobulin assembly
Abstract
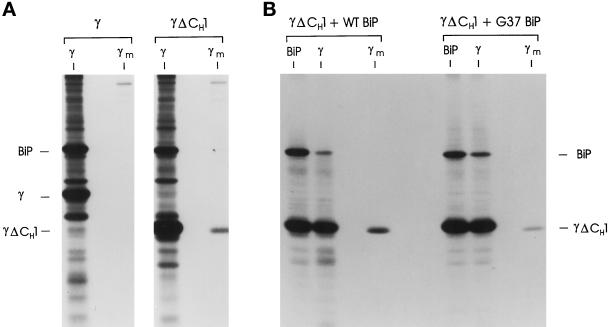
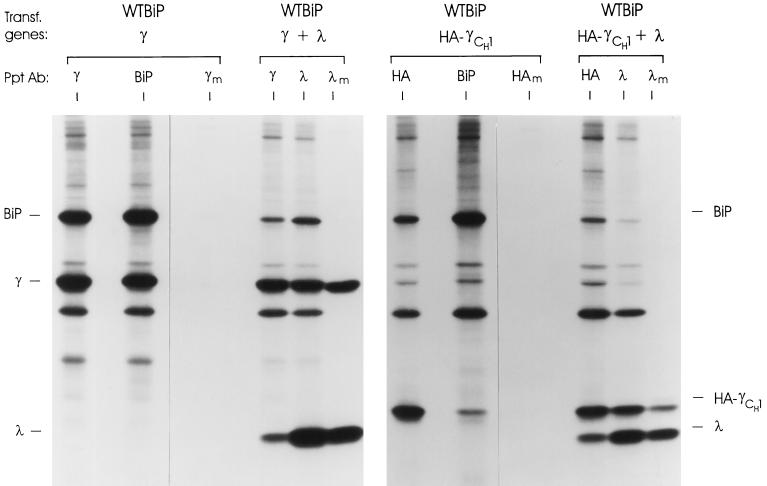
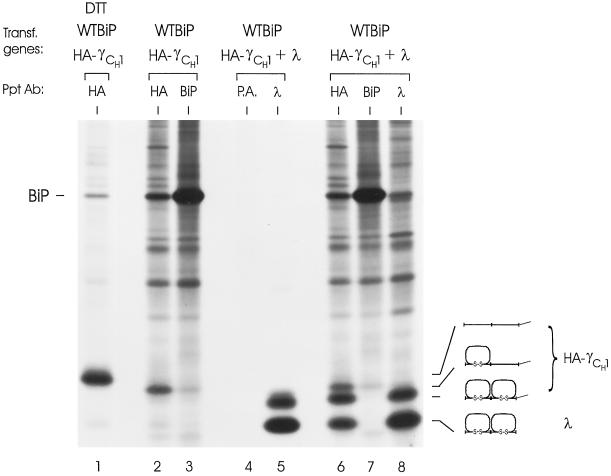
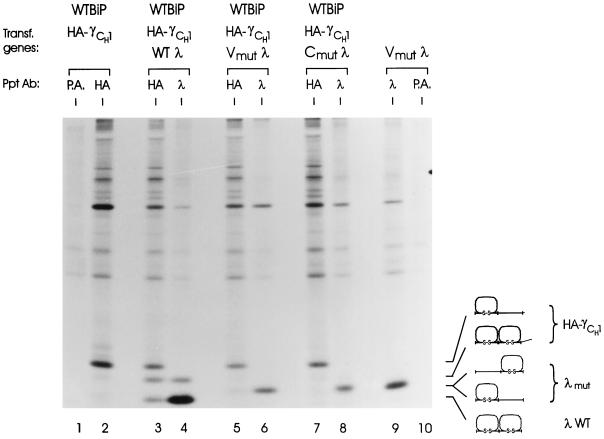
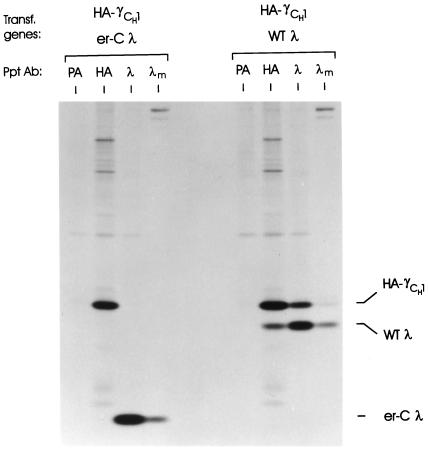
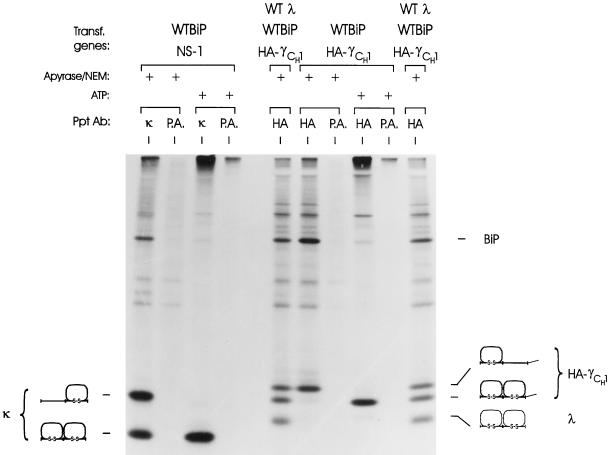
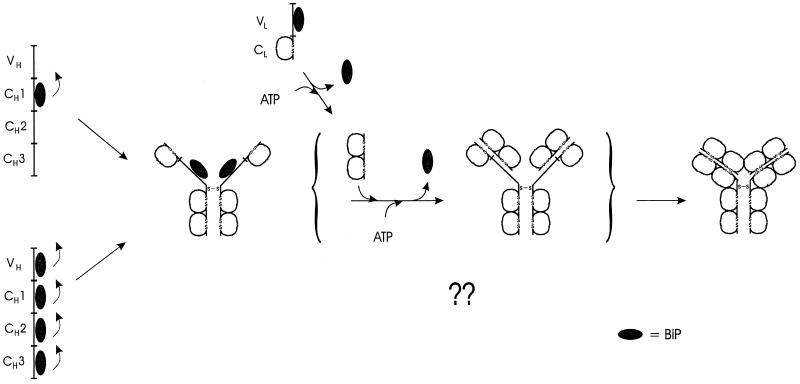
The immunoglobulin (Ig) molecule is composed of two identical heavy chains and two identical light chains (H2L2). Transport of this heteromeric complex is dependent on the correct assembly of the component parts, which is controlled, in part, by the association of incompletely assembled Ig heavy chains with the endoplasmic reticulum (ER) chaperone, BiP. Although other heavy chain-constant domains interact transiently with BiP, in the absence of light chain synthesis, BiP binds stably to the first constant domain (CH1) of the heavy chain, causing it to be retained in the ER. Using a simplified two-domain Ig heavy chain (VH-CH1), we have determined why BiP remains bound to free heavy chains and how light chains facilitate their transport. We found that in the absence of light chain expression, the CH1 domain neither folds nor forms its intradomain disulfide bond and therefore remains a substrate for BiP. In vivo, light chains are required to facilitate both the folding of the CH1 domain and the release of BiP. In contrast, the addition of ATP to isolated BiP-heavy chain complexes in vitro causes the release of BiP and allows the CH1 domain to fold in the absence of light chains. Therefore, light chains are not intrinsically essential for CH1 domain folding, but play a critical role in removing BiP from the CH1 domain, thereby allowing it to fold and Ig assembly to proceed. These data suggest that the assembly of multimeric protein complexes in the ER is not strictly dependent on the proper folding of individual subunits; rather, assembly can drive the complete folding of protein subunits.
Figures
Similar articles
-
Unassembled Ig heavy chains do not cycle from BiP in vivo but require light chains to trigger their release.Immunity. 2001 Jul;15(1):105-14. doi: 10.1016/s1074-7613(01)00163-7. Immunity. 2001. PMID: 11485742
-
Assembly and secretion of heavy chains that do not associate posttranslationally with immunoglobulin heavy chain-binding protein.J Cell Biol. 1987 Mar;104(3):761-7. doi: 10.1083/jcb.104.3.761. J Cell Biol. 1987. PMID: 3102505 Free PMC article.
-
The in vivo association of BiP with newly synthesized proteins is dependent on the rate and stability of folding and not simply on the presence of sequences that can bind to BiP.J Cell Biol. 1999 Jan 11;144(1):21-30. doi: 10.1083/jcb.144.1.21. J Cell Biol. 1999. PMID: 9885241 Free PMC article.
-
Biosynthesis, assembly and secretion of coagulation factor VIII.Blood Coagul Fibrinolysis. 1997 Dec;8 Suppl 2:S3-14. Blood Coagul Fibrinolysis. 1997. PMID: 9607108 Review.
-
ADP-ribosylation of the molecular chaperone GRP78/BiP.Mol Cell Biochem. 1994 Sep;138(1-2):141-8. doi: 10.1007/BF00928456. Mol Cell Biochem. 1994. PMID: 7898457 Review.
Cited by
-
Orchestration of secretory protein folding by ER chaperones.Biochim Biophys Acta. 2013 Nov;1833(11):2410-24. doi: 10.1016/j.bbamcr.2013.03.007. Epub 2013 Mar 15. Biochim Biophys Acta. 2013. PMID: 23507200 Free PMC article. Review.
-
Oligomerization of Hsp70: Current Perspectives on Regulation and Function.Front Mol Biosci. 2019 Sep 4;6:81. doi: 10.3389/fmolb.2019.00081. eCollection 2019. Front Mol Biosci. 2019. PMID: 31555664 Free PMC article. Review.
-
How antibodies fold.Trends Biochem Sci. 2010 Apr;35(4):189-98. doi: 10.1016/j.tibs.2009.11.005. Epub 2009 Dec 21. Trends Biochem Sci. 2010. PMID: 20022755 Free PMC article. Review.
-
Perturbation of the Relative Contribution of Molecular Chaperones in the Endoplasmic Reticulum.ACS Omega. 2020 Mar 24;5(13):7399-7405. doi: 10.1021/acsomega.9b04445. eCollection 2020 Apr 7. ACS Omega. 2020. PMID: 32280881 Free PMC article.
-
Antibody detection and kinetics of antibody production during early stages of immunization with hepatitis B virus vaccine.Clin Vaccine Immunol. 2007 Dec;14(12):1623-8. doi: 10.1128/CVI.00158-07. Epub 2007 Oct 10. Clin Vaccine Immunol. 2007. PMID: 17928429 Free PMC article.
References
-
- Amzel LM, Poljak RJ. Three-dimensional structure of immunoglobulins. Annu Rev Immunol. 1979;48:961–997. - PubMed
-
- Blond-Elguindi S, Cwirla SE, Dower WJ, Lipshutz RJ, Sprang SR, Sambrook JF, Gething MJ. Affinity panning of a library of peptides displayed on bacteriophages reveals the binding specificity of BiP. Cell. 1993;75:717–728. - PubMed
-
- Brightman SE, Blatch GL, Zetter BR. Isolation of a mouse cDNA encoding MTJ1, a new murine member of the DnaJ family of proteins. Gene. 1995;153:249–254. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
