

Mutagenesis of the Agrobacterium VirE2 single-stranded DNA-binding protein identifies regions required for self-association and interaction with VirE1 and a permissive site for hybrid protein construction
- PMID: 10400593
- PMCID: PMC93937
- DOI: 10.1128/JB.181.14.4342-4352.1999
Mutagenesis of the Agrobacterium VirE2 single-stranded DNA-binding protein identifies regions required for self-association and interaction with VirE1 and a permissive site for hybrid protein construction
Abstract
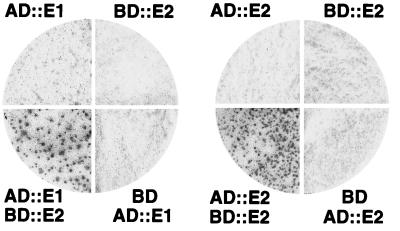
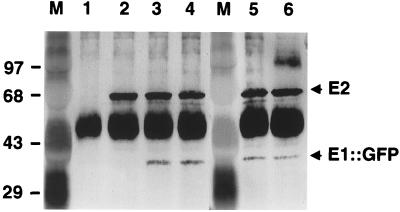
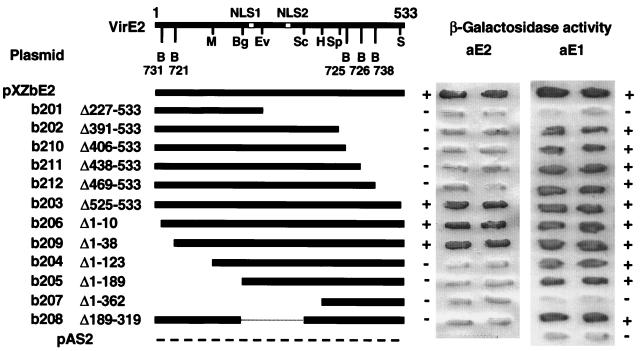
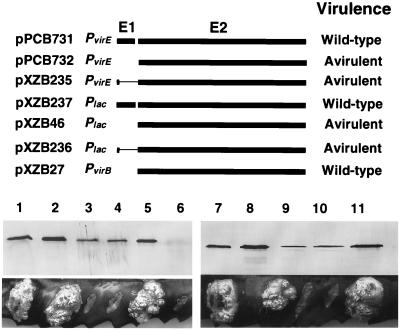
The VirE2 single-stranded DNA-binding protein (SSB) of Agrobacterium tumefaciens is required for delivery of T-DNA to the nuclei of susceptible plant cells. By yeast two-hybrid and immunoprecipitation analyses, VirE2 was shown to self-associate and to interact with VirE1. VirE2 mutants with small deletions or insertions of a 31-residue oligopeptide (i31) at the N or C terminus or with an i31 peptide insertion at Leu236 retained the capacity to form homomultimers. By contrast, VirE2 mutants with modifications outside a central region located between residues 320 and 390 retained the capacity to interact with VirE1. These findings suggest the tertiary structure of VirE2 is important for homomultimer formation whereas a central domain mediates formation of a complex with VirE1. The capacity of VirE2 mutants to interact with full-length VirE2 in the yeast Saccharomyces cerevisiae correlated with the abundance of the mutant proteins in A. tumefaciens, suggesting that VirE2 is stabilized by homomultimerization in the bacterium. We further characterized the promoter and N- and C-terminal sequence requirements for synthesis of functional VirE2. A PvirB::virE2 construct yielded functional VirE2 protein as defined by complementation of a virE2 null mutation. By contrast, PvirE or Plac promoter constructs yielded functional VirE2 only if virE1 was coexpressed with virE2. Deletion of 10 or 9 residues from the N or C terminus of VirE2, respectively, or addition of heterologous peptides or proteins to either terminus resulted in a loss of protein function. However, an i31 peptide insertion at Tyr39 had no effect on protein function as defined by the capacity of the mutant protein to (i) interact with native VirE2, (ii) interact with VirE1, (iii) accumulate at abundant levels in A. tumefaciens, and (iv) restore wild-type virulence to a virE2 null mutant. We propose that Tyr39 of VirE2 corresponds to a permissive site for insertion of heterologous peptides or proteins of interest for delivery across kingdom boundaries.
Figures



Similar articles
-
Functional domains of Agrobacterium tumefaciens single-stranded DNA-binding protein VirE2.J Bacteriol. 1997 Feb;179(4):1165-73. doi: 10.1128/jb.179.4.1165-1173.1997. J Bacteriol. 1997. PMID: 9023198 Free PMC article.
-
The Agrobacterium tumefaciens chaperone-like protein, VirE1, interacts with VirE2 at domains required for single-stranded DNA binding and cooperative interaction.J Bacteriol. 1999 Nov;181(21):6850-5. doi: 10.1128/JB.181.21.6850-6855.1999. J Bacteriol. 1999. PMID: 10542192 Free PMC article.
-
VirE1 is a specific molecular chaperone for the exported single-stranded-DNA-binding protein VirE2 in Agrobacterium.Mol Microbiol. 1999 Mar;31(6):1795-807. doi: 10.1046/j.1365-2958.1999.01316.x. Mol Microbiol. 1999. PMID: 10209751
-
The VirE2 protein of Agrobacterium tumefaciens: the Yin and Yang of T-DNA transfer.FEMS Microbiol Lett. 2003 Jun 6;223(1):1-6. doi: 10.1016/S0378-1097(03)00246-5. FEMS Microbiol Lett. 2003. PMID: 12798992 Review.
-
Agrobacterium tumefaciens and A. rhizogenes use different proteins to transport bacterial DNA into the plant cell nucleus.Microb Biotechnol. 2009 Jul;2(4):416-27. doi: 10.1111/j.1751-7915.2009.00104.x. Epub 2009 Apr 6. Microb Biotechnol. 2009. PMID: 21255274 Free PMC article. Review.
Cited by
-
Agrobacterium rhizogenes GALLS protein substitutes for Agrobacterium tumefaciens single-stranded DNA-binding protein VirE2.J Bacteriol. 2004 May;186(10):3065-77. doi: 10.1128/JB.186.10.3065-3077.2004. J Bacteriol. 2004. PMID: 15126468 Free PMC article.
-
The outs and ins of bacterial type IV secretion substrates.Trends Microbiol. 2003 Nov;11(11):527-35. doi: 10.1016/j.tim.2003.09.004. Trends Microbiol. 2003. PMID: 14607070 Free PMC article. Review.
-
A putative transmembrane leucine zipper of agrobacterium VirB10 is essential for t-pilus biogenesis but not type IV secretion.J Bacteriol. 2013 Jul;195(13):3022-34. doi: 10.1128/JB.00287-13. Epub 2013 Apr 26. J Bacteriol. 2013. PMID: 23625845 Free PMC article.
-
The All-Alpha Domains of Coupling Proteins from the Agrobacterium tumefaciens VirB/VirD4 and Enterococcus faecalis pCF10-Encoded Type IV Secretion Systems Confer Specificity to Binding of Cognate DNA Substrates.J Bacteriol. 2015 Jul;197(14):2335-49. doi: 10.1128/JB.00189-15. Epub 2015 May 4. J Bacteriol. 2015. PMID: 25939830 Free PMC article.
-
DNA substrate-induced activation of the Agrobacterium VirB/VirD4 type IV secretion system.J Bacteriol. 2013 Jun;195(11):2691-704. doi: 10.1128/JB.00114-13. Epub 2013 Apr 5. J Bacteriol. 2013. PMID: 23564169 Free PMC article.
References
-
- Anderson D M, Schneewind O. Type III machines of gram-negative pathogens: injecting virulence factors into host cells and more. Curr Opin Microbiol. 1999;2:18–24. . (Review.) - PubMed
-
- Bai C, Elledge S J. Gene identification using the yeast two-hybrid system. Methods Enzymol. 1996;273:331–347. - PubMed
-
- Bartel P L, Fields S. The yeast two-hybrid system. Oxford, United Kingdom: Oxford University Press; 1997.
-
- Beijersbergen A, Dulk-Ras A D, Schilperoort R A, Hooykaas P J J. Conjugative transfer by the virulence system of Agrobacterium tumefaciens. Science. 1992;256:1324–1327. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
