

Identification and complementation of frameshift mutations associated with loss of cytadherence in Mycoplasma pneumoniae
- PMID: 10400600
- PMCID: PMC93944
- DOI: 10.1128/JB.181.14.4404-4410.1999
Identification and complementation of frameshift mutations associated with loss of cytadherence in Mycoplasma pneumoniae
Abstract
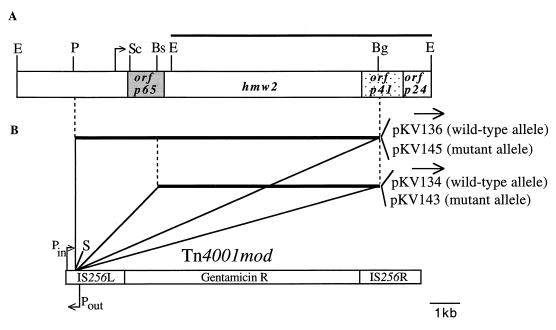
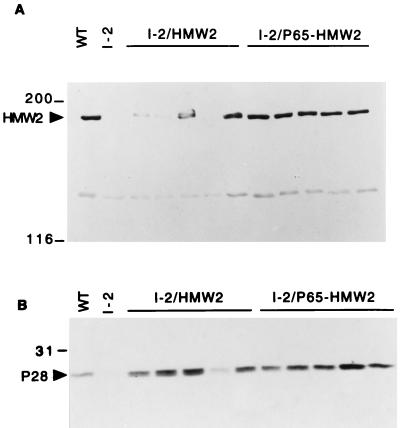
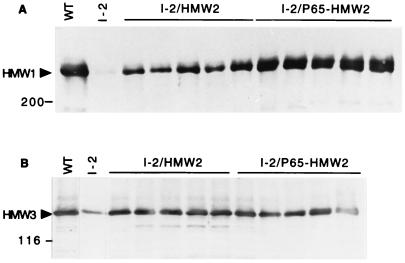
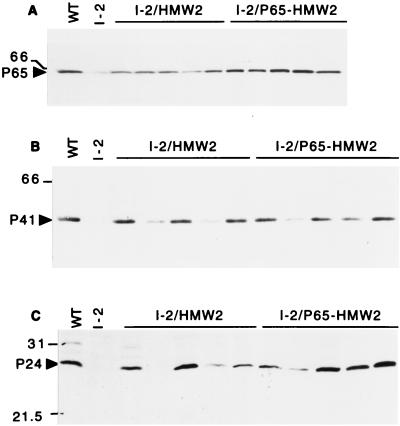
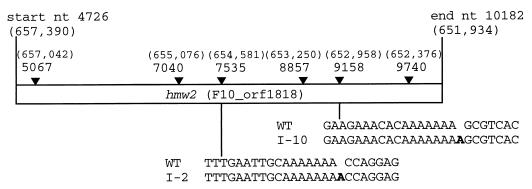
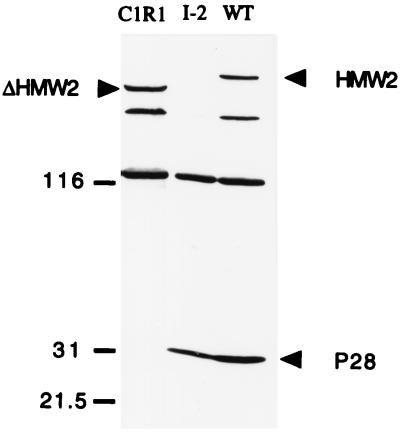
Mycoplasma pneumoniae cytadherence is mediated by a specialized, polar attachment organelle. Certain spontaneously arising cytadherence mutants (designated class I) lack HMW2, fail to localize the adhesin protein P1 to the attachment organelle, and exhibit accelerated turnover of proteins HMW1, HMW3, and P65. Insertional inactivation of hmw2 by Tn4001 results in a phenotype nearly identical to that of the class I mutants, suggesting that the latter may result from a defect in hmw2. In this study, the recombinant wild-type hmw2 allele successfully complemented a class I mutant when introduced by transposon delivery. Synthesis of recombinant HMW2 at wild-type levels resulted in reacquisition of hemadsorption and normal levels of HMW1, HMW3, and P65. Low-level production of HMW2 in some transformants resulted in only an intermediate capacity to hemadsorb. Furthermore, full restoration of HMW1 and P65, but not that of HMW3, was directly proportional to the amount of recombinant HMW2 produced, reflecting the importance of proper stoichiometry for certain cytadherence-associated proteins. The recombinant class I hmw2 allele did not restore cytadherence, consistent with a defect in hmw2 in this mutant. A frameshift was discovered in different oligoadenine tracts in hmw2 from two independent class I mutants. Finally, protein P28 is thought to be the product of internal translation initiation in hmw2. A transposon excision-deletion mutant produced a truncated HMW2 but no P28, consistent with this conclusion. However, this deletion mutant was hemadsorption positive, indicating that P28 may not be required for cytadherence.
Figures
Similar articles
-
Deletion analysis identifies key functional domains of the cytadherence-associated protein HMW2 of Mycoplasma pneumoniae.Mol Microbiol. 2003 Dec;50(5):1507-16. doi: 10.1046/j.1365-2958.2003.03807.x. Mol Microbiol. 2003. PMID: 14651634
-
Transposon mutagenesis reinforces the correlation between Mycoplasma pneumoniae cytoskeletal protein HMW2 and cytadherence.J Bacteriol. 1997 Apr;179(8):2668-77. doi: 10.1128/jb.179.8.2668-2677.1997. J Bacteriol. 1997. PMID: 9098066 Free PMC article.
-
Attachment organelle formation represented by localization of cytadherence proteins and formation of the electron-dense core in wild-type and mutant strains of Mycoplasma pneumoniae.J Bacteriol. 2003 Feb;185(3):1082-91. doi: 10.1128/JB.185.3.1082-1091.2003. J Bacteriol. 2003. PMID: 12533484 Free PMC article.
-
Mycoplasma pneumoniae cytadherence: unravelling the tie that binds.Mol Microbiol. 1996 Apr;20(2):247-53. doi: 10.1111/j.1365-2958.1996.tb02613.x. Mol Microbiol. 1996. PMID: 8733224 Review.
-
Mycoplasma pneumoniae cytadherence: organization and assembly of the attachment organelle.Trends Microbiol. 1998 Jan;6(1):15-8. doi: 10.1016/S0966-842X(97)01168-2. Trends Microbiol. 1998. PMID: 9481818 Review.
Cited by
-
Domain analysis of protein P30 in Mycoplasma pneumoniae cytadherence and gliding motility.J Bacteriol. 2011 Apr;193(7):1726-33. doi: 10.1128/JB.01228-10. Epub 2011 Jan 21. J Bacteriol. 2011. PMID: 21257768 Free PMC article.
-
Molecular ruler of the attachment organelle in Mycoplasma pneumoniae.PLoS Pathog. 2021 Jun 10;17(6):e1009621. doi: 10.1371/journal.ppat.1009621. eCollection 2021 Jun. PLoS Pathog. 2021. PMID: 34111235 Free PMC article.
-
Protein P200 is dispensable for Mycoplasma pneumoniae hemadsorption but not gliding motility or colonization of differentiated bronchial epithelium.Infect Immun. 2007 Jan;75(1):518-22. doi: 10.1128/IAI.01344-06. Epub 2006 Oct 16. Infect Immun. 2007. PMID: 17043103 Free PMC article.
-
Stability of Mycoplasma pneumoniae cytadherence-accessory protein HMW1 correlates with its association with the triton shell.J Bacteriol. 2001 Jun;183(12):3680-8. doi: 10.1128/JB.183.12.3680-3688.2001. J Bacteriol. 2001. PMID: 11371532 Free PMC article.
-
Visualization of the attachment organelle and cytadherence proteins of Mycoplasma pneumoniae by immunofluorescence microscopy.J Bacteriol. 2001 Mar;183(5):1621-30. doi: 10.1128/JB.183.5.1621-1630.2001. J Bacteriol. 2001. PMID: 11160093 Free PMC article.
References
-
- Dirksen L B, Proft T, Hilbert H, Plagens H, Herrmann R, Krause D C. Nucleotide sequence analysis and characterization of the hmw gene cluster of Mycoplasma pneumoniae. Gene. 1996;171:19–25. - PubMed
-
- Dovc P, Bencina D, Milosevic T, Berlic M D-V, Narat M. Presented at the 12th Congress of the International Organization for Mycoplasmology, Sydney, Australia, July 22 to 28. 1998. Localized changes in Mycoplasma gallisepticum pMGA gene associated with antigenic variants.
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
