

Structure of adenovirus complexed with its internalization receptor, alphavbeta5 integrin
- PMID: 10400774
- PMCID: PMC112761
- DOI: 10.1128/JVI.73.8.6759-6768.1999
Structure of adenovirus complexed with its internalization receptor, alphavbeta5 integrin
Abstract
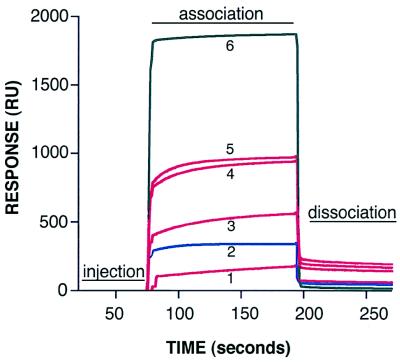
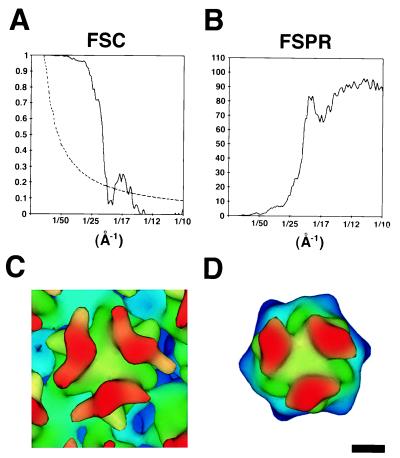
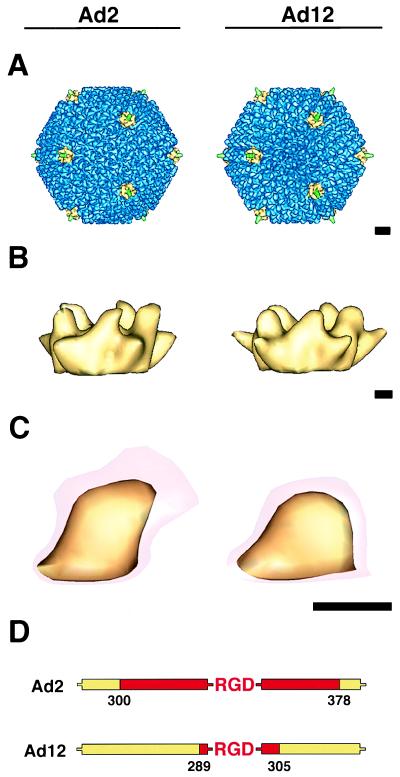
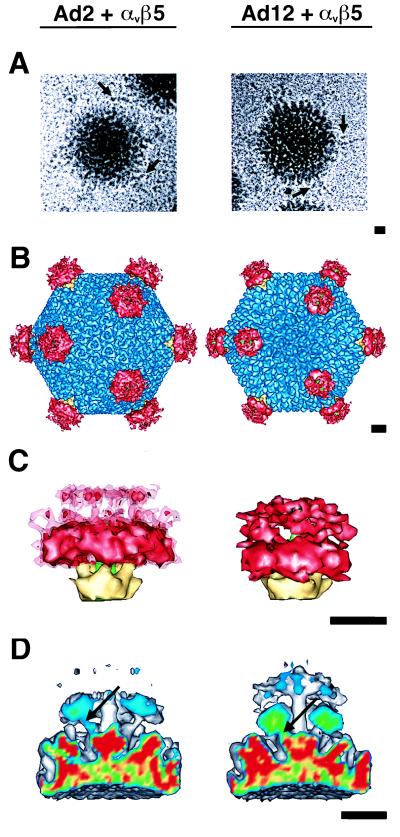
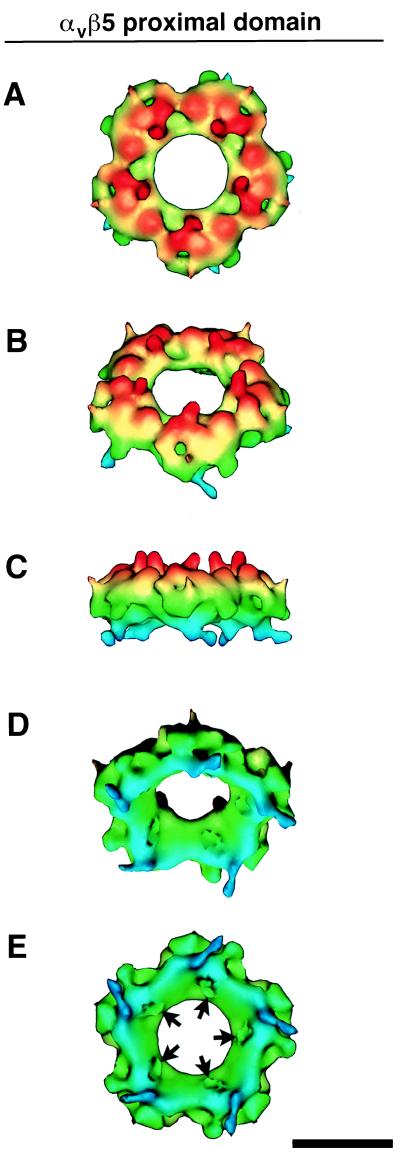
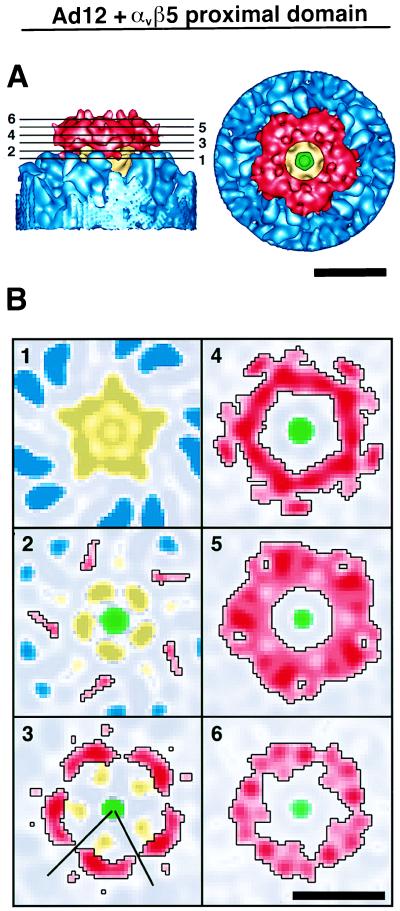
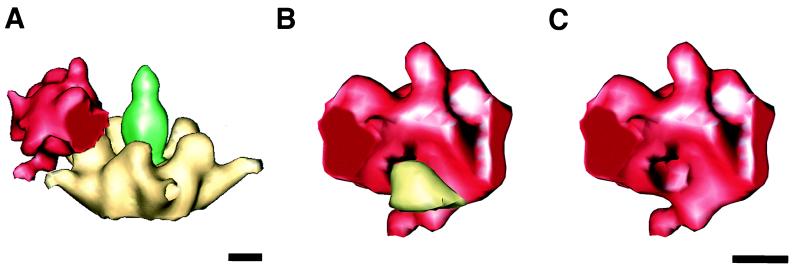
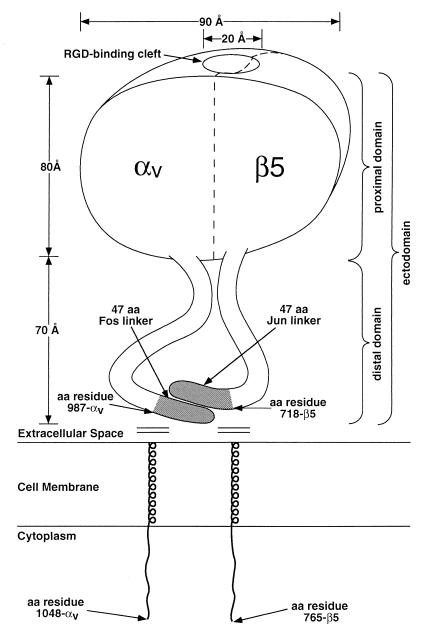
The three-dimensional structure of soluble recombinant integrin alphavbeta5 bound to human adenovirus types 2 and 12 (Ad2 and -12) has been determined at approximately 21-A resolution by cryoelectron microscopy (cryo-EM). The alphavbeta5 integrin is known to promote Ad cell entry. Cryo-EM has shown that the integrin-binding RGD (Arg-Gly-Asp) protrusion of the Ad2 penton base protein is highly mobile (P. L. Stewart, C. Y. Chiu, S. Huang, T. Muir, Y. Zhao, B. Chait, P. Mathias, and G. R. Nemerow, EMBO J. 16:1189-1198, 1997). Sequence analysis indicated that the Ad12 RGD surface loop is shorter than that of Ad2 and probably less flexible, hence more suitable for structural characterization of the Ad-integrin complex. The cryo-EM structures of the two virus-receptor complexes revealed a ring of integrin density above the penton base of each virus serotype. As expected, the integrin density in the Ad2 complex was diffuse while that in the Ad12 complex was better defined. The integrin consists of two discrete subdomains, a globular domain with an RGD-binding cleft approximately 20 A in diameter and a distal domain with extended, flexible tails. Kinetic analysis of Ad2 interactions with alphavbeta5 indicated approximately 4.2 integrin molecules bound per penton base at close to saturation. These results suggest that the precise spatial arrangement of five RGD protrusions on the penton base promotes integrin clustering and the signaling events required for virus internalization.
Figures
References
-
- Acharya R, Fry E, Stuart D, Fox G, Rowlands D, Brown F. The three-dimensional structure of foot-and-mouth disease virus at 2.9 Å resolution. Nature. 1989;337:709–716. - PubMed
-
- Adler M, Lazarus R A, Dennis M S, Wagner G. Solution structure of kistrin, a potent platelet aggregation inhibitor and GP IIb-IIIa antagonist. Science. 1991;253:445–448. - PubMed
-
- Adrian M, Dubochet J, Lepault J, McDowall A W. Cryo-electron microscopy of viruses. Nature. 1984;308:32–36. - PubMed
-
- Akke M, Liu J, Cavanagh J, Erickson H P, Palmer A G I. Pervasive conformational fluctuations on microsecond time scales in a fibronectin type III domain. Nat Struct Biol. 1998;5:55–59. - PubMed
-
- Altschul S F, Gish W, Miller W, Myers E W, Lipman D J. Basic local alignment search tool. J Mol Biol. 1990;215:403–410. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
