

Long-term infection and transformation of dermal microvascular endothelial cells by human herpesvirus 8
- PMID: 10400787
- PMCID: PMC112774
- DOI: 10.1128/JVI.73.8.6892-6902.1999
Long-term infection and transformation of dermal microvascular endothelial cells by human herpesvirus 8
Abstract
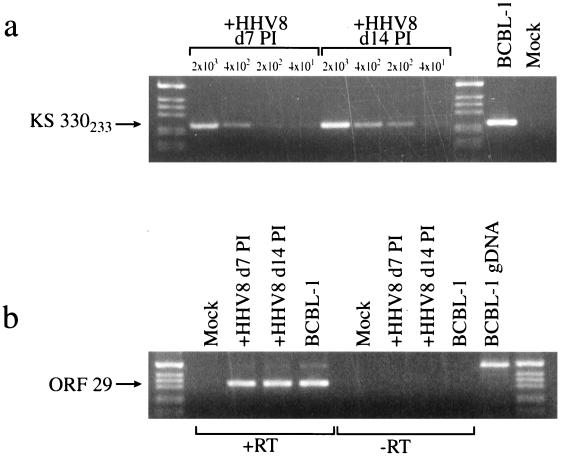
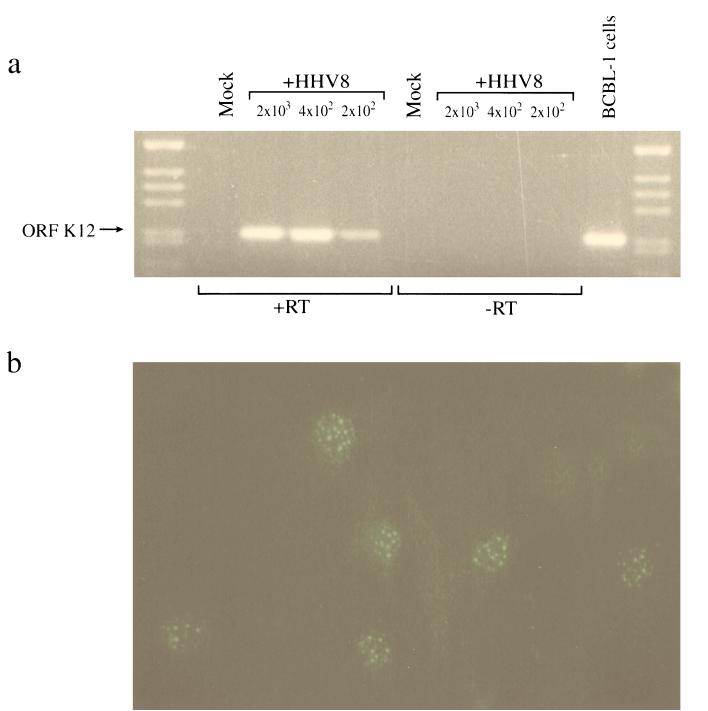
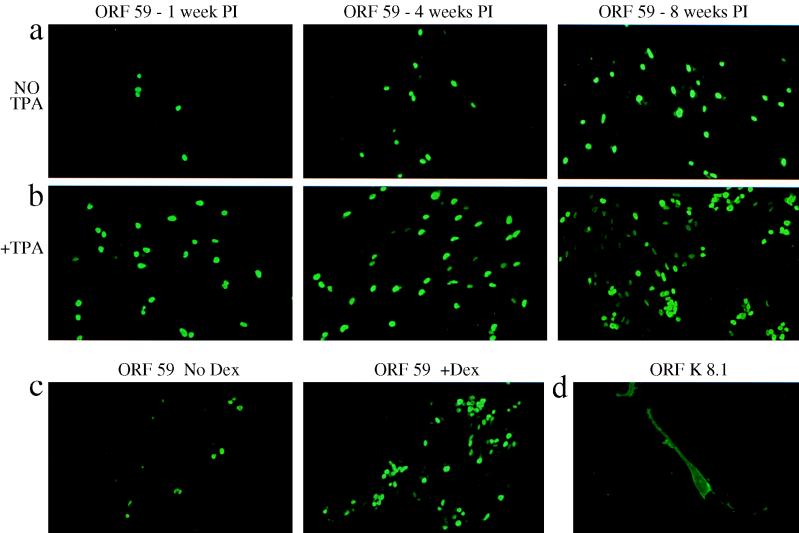
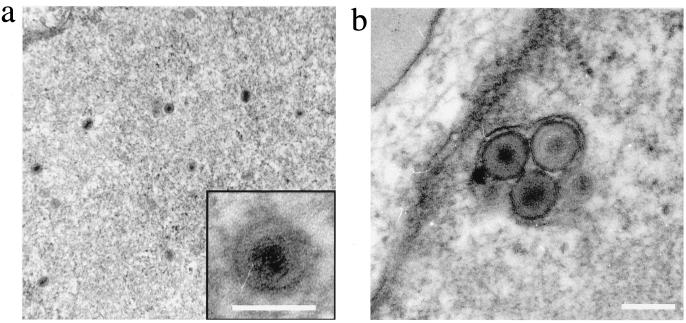
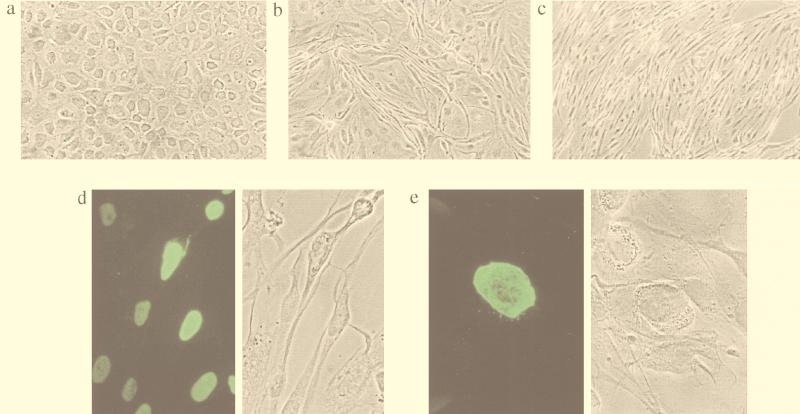
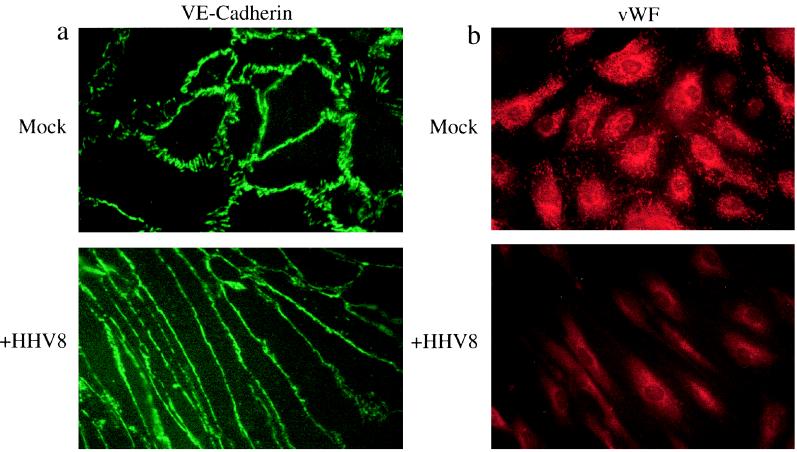
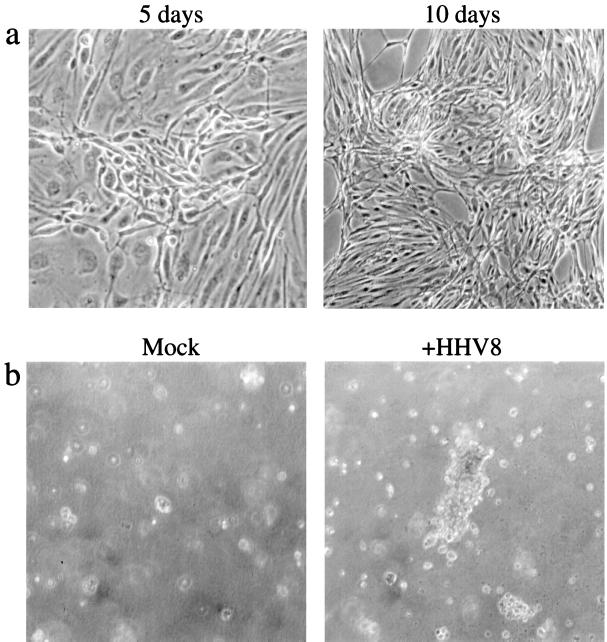
Human herpesvirus 8 (HHV8) infects Kaposi's sarcoma (KS) spindle cells in situ, as well as the lesional endothelial cells considered to be spindle cell precursors. The HHV8 genome contains several oncogenes, suggesting that infection of endothelial and spindle cells could induce cellular transformation and tumorigenesis and promote the formation of KS lesions. To investigate the potential of HHV8 infection of endothelial cells to contribute to the development of KS, we have developed an in vitro model utilizing dermal microvascular endothelial cells that support significant HHV8 infection. In contrast to existing in vitro systems used to study HHV8 pathogenesis, the majority of dermal endothelial cells are infected with HHV8 and the viral genome is maintained indefinitely. Infection is predominantly latent, with a small percentage of cells supporting lytic replication, and latency is responsive to lytic induction stimuli. Infected endothelial cells develop a spindle shape resembling that of KS lesional cells and show characteristics of a transformed phenotype, including loss of contact inhibition and acquisition of anchorage-independent growth. These results describe a relevant model system in which to study virus-host interactions in vitro and demonstrate the ability of HHV8 to induce phenotypic changes in infected endothelial cells that resemble characteristics of KS spindle cells in vivo. Thus, our results are consistent with a direct role for HHV8 in the pathogenesis of KS.
Figures
Similar articles
-
Spindle cell conversion by Kaposi's sarcoma-associated herpesvirus: formation of colonies and plaques with mixed lytic and latent gene expression in infected primary dermal microvascular endothelial cell cultures.J Virol. 2001 Jun;75(12):5614-26. doi: 10.1128/JVI.75.12.5614-5626.2001. J Virol. 2001. PMID: 11356969 Free PMC article.
-
De novo infection and serial transmission of Kaposi's sarcoma-associated herpesvirus in cultured endothelial cells.J Virol. 2002 Mar;76(5):2440-8. doi: 10.1128/jvi.76.5.2440-2448.2002. J Virol. 2002. PMID: 11836422 Free PMC article.
-
[Kaposi's disease and HHV8: a new model of virus-induced tumorigenesis].Pathol Biol (Paris). 2000 Jul;48(6):529-32. Pathol Biol (Paris). 2000. PMID: 10965529 French.
-
Biology of Kaposi's sarcoma.Eur J Cancer. 2001 Jul;37(10):1251-69. doi: 10.1016/s0959-8049(01)00121-6. Eur J Cancer. 2001. PMID: 11423257 Review.
-
Endothelial cell- and lymphocyte-based in vitro systems for understanding KSHV biology.Curr Top Microbiol Immunol. 2007;312:211-44. doi: 10.1007/978-3-540-34344-8_8. Curr Top Microbiol Immunol. 2007. PMID: 17089799 Review.
Cited by
-
Infection and transmission dynamics of rKSHV.219 in primary endothelial cells.J Virol Methods. 2013 Oct;193(1):251-9. doi: 10.1016/j.jviromet.2013.06.001. Epub 2013 Jun 10. J Virol Methods. 2013. PMID: 23764419 Free PMC article.
-
Envelope glycoprotein gB of Kaposi's sarcoma-associated herpesvirus is essential for egress from infected cells.J Virol. 2005 Sep;79(17):10952-67. doi: 10.1128/JVI.79.17.10952-10967.2005. J Virol. 2005. PMID: 16103147 Free PMC article.
-
Effective human herpesvirus 8 infection of human umbilical vein endothelial cells by cell-mediated transmission.J Virol. 2001 Aug;75(16):7717-22. doi: 10.1128/JVI.75.16.7717-7722.2001. J Virol. 2001. PMID: 11462044 Free PMC article.
-
Inflammatory cytokines inhibit Kaposi's sarcoma-associated herpesvirus lytic gene transcription in in vitro-infected endothelial cells.J Virol. 2004 Mar;78(5):2591-6. doi: 10.1128/jvi.78.5.2591-2596.2004. J Virol. 2004. PMID: 14963163 Free PMC article.
-
Patterns of gene expression and a transactivation function exhibited by the vGCR (ORF74) chemokine receptor protein of Kaposi's sarcoma-associated herpesvirus.J Virol. 2002 Apr;76(7):3421-39. doi: 10.1128/jvi.76.7.3421-3439.2002. J Virol. 2002. PMID: 11884567 Free PMC article.
References
-
- Aluigi M G, Albini A, Carlone S, Repetto L, De Marchi R, Icardi A, Moro M, Noonan D, Benelli R. KSHV sequences in biopsies and cultured spindle cells of epidemic, iatrogenic and Mediterranean forms of Kaposi’s sarcoma. Res Virol. 1996;147:267–275. - PubMed
-
- Ambroziak J A, Blackbourn D J, Herndier B G, Glougau R G, Gullet J H, McDonald A R, Lennette E T, Levy J A. Herpes-like sequences in HIV-infected and uninfected Kaposi’s sarcoma patients. Science. 1995;268:582–583. - PubMed
-
- Arvanitakis L, Mesri E A, Nador R G, Said J W, Asch A S, Knowles D M, Cesarman E. Establishment and characterization of a primary effusion (body cavity-based) lymphoma cell line (BC-3) harboring Kaposi’s sarcoma-associated herpesvirus (KSHV/HHV-8) in the absence of Epstein-Barr virus. Blood. 1996;88:2648–2654. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
