

doi: 10.1128/JVI.73.8.7070-7076.1999.
Resistance of ribosomal protein mRNA translation to protein synthesis shutoff induced by poliovirus
Affiliations
- PMID: 10400812
- PMCID: PMC112799
- DOI: 10.1128/JVI.73.8.7070-7076.1999
Item in Clipboard
Resistance of ribosomal protein mRNA translation to protein synthesis shutoff induced by poliovirus
J Virol.
1999 Aug.
Abstract
Poliovirus infection induces an overall inhibition of host protein synthesis, although some mRNAs continue to be translated, suggesting different translation requirements for cellular mRNAs. It is known that ribosomal protein mRNAs are translationally regulated and that the phosphorylation of ribosomal protein S6 is involved in the regulation. Here, we report that the translation of ribosomal protein mRNAs resists poliovirus infection and correlates with an increase in p70(s6k) activity and phosphorylation of ribosomal protein S6.
Figures
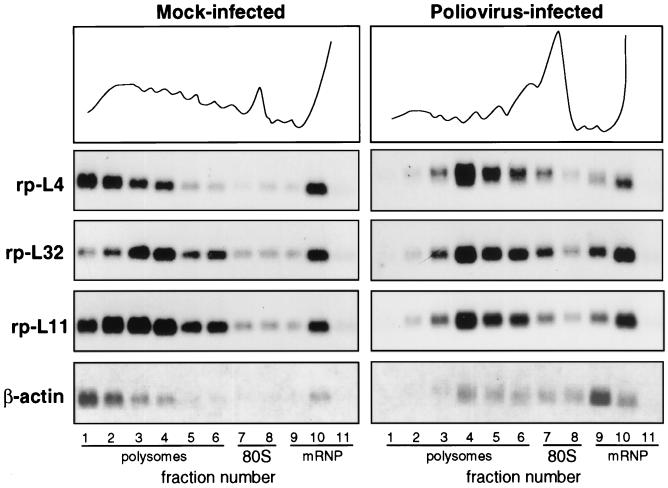
Polysome-mRNP distribution of mRNA in mock-infected and poliovirus-infected cells. HEp-2 cells were grown at 37°C in Eagle’s minimum essential medium supplemented with 10% fetal calf serum. When cells reached 80% confluence, the Mahoney strain of poliovirus type 1 was added at a multiplicity of infection of 50 PFU per cell. A 9-cm-diameter plate culture of mock-infected and infected cells was lysed at 4 h postinfection (10 mM NaCl, 10 mM MgCl2, 1% Triton X-100, 1% Na-deoxycholate, 1 mM dithiothreitol, 10 mM Tris-HCl, pH 7.4) to prepare cytoplasmic extracts (32). Cycloheximide, often used to prevent polysome runoff, was not added prior to extract preparation, as this drug sometime creates problems (38). Polysome protection can also be obtained by quickly preparing the extract and loading the sample on the gradient under strict temperature control. The extracts were separated on 15 to 50% sucrose gradients in gradient buffer (0.1 M NaCl, 10 mM MgCl2, 30 mM Tris-HCl, pH 7). Gradient fractions were collected while the optical density profile at 260 nm was monitored (top), and the RNA was prepared by protease K-SDS-phenol extraction (32) of the fractions. The RNAs from equal volumes of mock-infected and infected gradient fractions were loaded onto two gels and analyzed by Northern blotting and autoradiography as previously described (32). Each filter was subsequently hybridized to probes for rp-L4 (2), rp-L32 (10), rp-L11 (12), and β-actin (6) mRNAs to obtain a reliable comparison of the distribution of the various RNAs along the same gradient. Probes were prepared by the random primer technique.
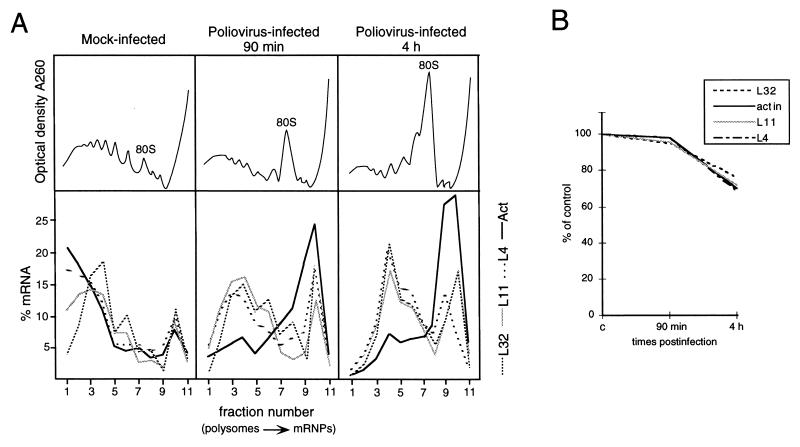
Time course of polysome-mRNP distribution and accumulation of mRNAs during poliovirus infection. (A) Mock-infected and poliovirus-infected cells were processed at the times indicated as described in the legend to Fig. 1. Northern blots were subsequently hybridized with rp-L4, rp-L11, rp-L32, and β-actin probes. Measurement of radioactivity, reported as the percentage of mRNA in each fraction, was done by PhosphorImager (Molecular Dynamics) analysis. The optical density profiles of the sucrose gradients were monitored at 260 nm (top), and the positions of the 80S monomers are indicated. These experiments were performed at least three times, and the results were consistently similar. (B) One tenth of each extract, prepared at the indicated time postinfection, was taken before gradient loading. The RNA was extracted, and equivalent amounts were analyzed by Northern blotting as described in the legend to Fig. 1. The filters were subsequently hybridized to rp-L4, rp-L32, rp-L11, β-actin, and 5S RNA probes (46). Measurement of radioactivity was done by PhosphorImager (Molecular Dynamics) analysis, and the values obtained were normalized to the signal of the 5S rRNA probe. The level of each mRNA is expressed as a percentage of the amount measured in mock-infected cells (c).
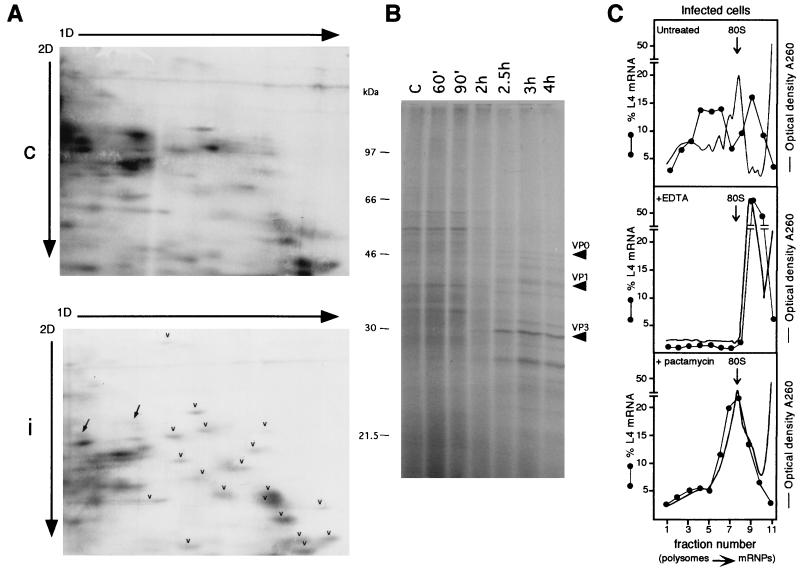
Analysis of the activity of polysome-associated RNA in infected cells by metabolic labeling of proteins and by EDTA and pactamycin treatment. (A) HEp-2 cells were grown and infected as described in the legend to Fig. 1. At 90 min postinfection, mock-infected and infected cells were incubated for 30 min with [35S]methionine-[35S]cysteine (Pro-mix; Amersham; >1,000 Ci/mmol) at a concentration of 0.1 mCi/ml. Cells were harvested in phosphate-buffered saline and homogenized in 0.5 ml of ice-cold rp buffer (0.1 M NaCl, 1 mM MgCl2, 10 mM HEPES, pH 7.5), acid extracted (58), and processed by the 2D gel electrophoresis method optimized to resolve r proteins, with the exception that the second-dimension (2D) gel was 13% polyacrylamide (58). A 200-μg sample of r proteins purified from HEp-2 ribosomes (58) was added as markers to an equal amount of control (c) or infected-cell (i) protein extract (200 μg). Gels were fluorographed and exposed to X-ray film for the same time. Arrowheads point to some r proteins, and arrows point to non-r proteins. 1D, first dimension. (B) HEp-2 cells, grown and infected as indicated above, were labeled with Pro-mix (40 μCi/ml) for 10 min at the indicated times after infection. Cells were lysed as described above, and 5 μg of each extract was loaded onto an SDS–12.5% PAGE gel and autoradiographed. Arrows point to viral proteins. Lane C, control. (C) Untreated infected cells (top) were lysed at 4 h postinfection and analyzed on a sucrose gradient as described in the legend to Fig. 1. For the EDTA treatment (middle), cytoplasmic extracts, brought to a concentration of 50 mM EDTA, pH 7.4, were incubated in ice for 5 min, loaded onto sucrose gradients containing 10 mM EDTA instead of magnesium, and analyzed as described in the legend to Fig. 1. To test the effect of pactamycin (bottom), cells were incubated at 3.5 h after infection with 30 ng of pactamycin per ml for 30 min, thus reaching the 4-h infection time of untreated cells, and then processed as described in the legend to Fig. 1. When the gradient fractions were collected, the optical density at 260 nm was monitored. The profiles are shown as a continuous line, and the 80S monomers are indicated by the arrows. Northern blots were hybridized with an rp-L4 probe. Measurement of radioactivity, reported as a percentage of the mRNA in each fraction, was done by PhosphorImager (Molecular Dynamics) analysis.
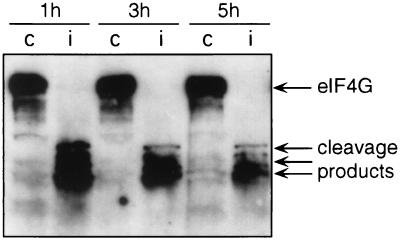
eIF4G cleavage in poliovirus-infected cells. Cytoplasmic extracts from mock-infected (lanes c) and poliovirus-infected (lanes i) cells were prepared at the times indicated as described in the legend to Fig. 1. Aliquots of the cytoplasmic extracts were precipitated with acetone for protein analysis by SDS-PAGE and Western blotting using the anti-eIF4G antibody as previously described (18).
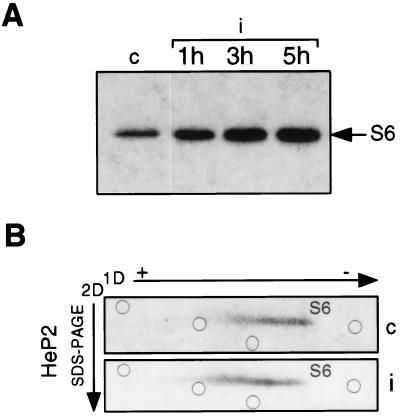
p70S6k activity and S6 phosphorylation during poliovirus infection. (A) S100 cytoplasmic extracts from mock-infected (lane c) and poliovirus-infected (i) cells were prepared at the times indicated in the presence of a phosphatase inhibitor as previously described (40). Protein concentration was determined by the Bio-Rad Protein Assay kit. Eight-microgram extract samples were used for immunoprecipitation of p70S6k with M6 antibody, and the immunocomplex was assayed for in vitro kinase activity in the presence of 40S ribosomes at an A260 of 0.45 U as previously described (16). Proteins were separated by SDS–PAGE and autoradiographed. Phosphorylated S6 protein is indicated by the arrow. (B) Mock-infected and poliovirus-infected cells (c and i, respectively) were labeled at 2.5 h after infection in a phosphate-free medium with 20 μCi of [32P]orthophosphate per ml for 90 min. Ribosomes from labeled cells were purified, and ribosomal proteins were extracted in the presence of 40 mM β-glycerophosphate (58). Proteins were analyzed by 2D electrophoresis as previously described (33), with the exception that the second dimension (2D) was an SDS–15% PAGE gel. A 200-μg sample of unlabeled purified HEp-2 ribosomal proteins was added to each sample before gel loading for Coomassie staining and used as position markers in the gel. The positions of stained ribosomal proteins in the S6 area are indicated by open circles. To standardize 2D gel electrophoresis conditions and to map the positions of S6 phosphorylated forms, preliminary experiments were carried out by loading gels with purified 40S subunits phosphorylated in in vitro kinase assays by extracts from quiescent and serum-stimulated Swiss 3T3 cells (16). The level of S6 phosphorylation by quiescent cell extracts was very low and increased 20-fold after serum stimulation. Accordingly, an evident shift in migration of S6 in the 2D gel was observed (data not shown). 1D, first dimension.
Similar articles
-
S6 phosphorylation-independent pathways regulate translation of 5'-terminal oligopyrimidine tract-containing mRNAs in differentiating hematopoietic cells.Nucleic Acids Res. 2002 May 1;30(9):1919-28. doi: 10.1093/nar/30.9.1919. Nucleic Acids Res. 2002. PMID: 11972328 Free PMC article.
-
Targeted disruption of p70(s6k) defines its role in protein synthesis and rapamycin sensitivity.Proc Natl Acad Sci U S A. 1998 Apr 28;95(9):5033-8. doi: 10.1073/pnas.95.9.5033. Proc Natl Acad Sci U S A. 1998. PMID: 9560223 Free PMC article.
-
Phosphorylation of ribosomal protein S6 by cAMP-dependent protein kinase and mitogen-stimulated S6 kinase differentially alters translation of globin mRNA.J Biol Chem. 1987 Mar 15;262(8):3518-23. J Biol Chem. 1987. PMID: 3818653
-
Regulation of global and specific mRNA translation by amino acids.J Nutr. 2002 May;132(5):883-6. doi: 10.1093/jn/132.5.883. J Nutr. 2002. PMID: 11983807 Review.
-
Ribosomal protein S6 kinase from TOP mRNAs to cell size.Prog Mol Biol Transl Sci. 2009;90:109-53. doi: 10.1016/S1877-1173(09)90003-5. Epub 2009 Oct 27. Prog Mol Biol Transl Sci. 2009. PMID: 20374740 Review.
Cited by
-
Unveiling the role of hexon-associated host proteins in fowl adenovirus serotype 4 replication.Front Vet Sci. 2025 Jun 3;12:1562872. doi: 10.3389/fvets.2025.1562872. eCollection 2025. Front Vet Sci. 2025. PMID: 40530025 Free PMC article.
-
Genome-Scale CRISPR Screening Reveals Host Factors Required for Ribosome Formation and Viral Replication.mBio. 2023 Apr 25;14(2):e0012723. doi: 10.1128/mbio.00127-23. Epub 2023 Feb 21. mBio. 2023. PMID: 36809113 Free PMC article.
-
Selective translation of eukaryotic mRNAs: functional molecular analysis of GRSF-1, a positive regulator of influenza virus protein synthesis.J Virol. 2002 Oct;76(20):10417-26. doi: 10.1128/jvi.76.20.10417-10426.2002. J Virol. 2002. PMID: 12239318 Free PMC article.
-
Ribosomal protein S6 associates with alphavirus nonstructural protein 2 and mediates expression from alphavirus messages.J Virol. 2006 Aug;80(15):7729-39. doi: 10.1128/JVI.00425-06. J Virol. 2006. PMID: 16840351 Free PMC article.
-
Regulation of Ribosomal Proteins on Viral Infection.Cells. 2019 May 27;8(5):508. doi: 10.3390/cells8050508. Cells. 2019. PMID: 31137833 Free PMC article. Review.
References
-
- Amaldi F, Pierandrei-Amaldi P. TOP genes: a translationally controlled class of genes including those coding for ribosomal proteins. In: Jeanteur P, editor. Cytoplasmic fate of eukaryotic mRNA. Vol. 18. Heidelberg, Germany: Springer-Verlag; 1997. pp. 1–17. - PubMed
-
- Bagni C, Mariottini P, Annesi F, Amaldi F. Human ribosomal protein L4: cloning and sequencing of the cDNA and primary structure of the protein. Biochim Biophys Acta. 1993;1216:475–478. - PubMed
-
- Brown E J, Schreiber S L. A signaling pathway to translational control. Cell. 1996;86:517–520. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
