

Intracellular localization of proteasomal degradation of a viral antigen
- PMID: 10402464
- PMCID: PMC2199725
- DOI: 10.1083/jcb.146.1.113
Intracellular localization of proteasomal degradation of a viral antigen
Abstract
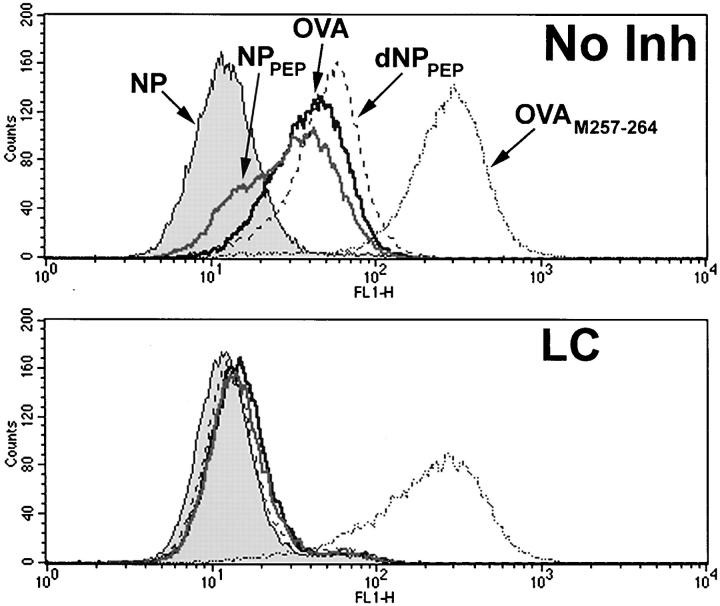
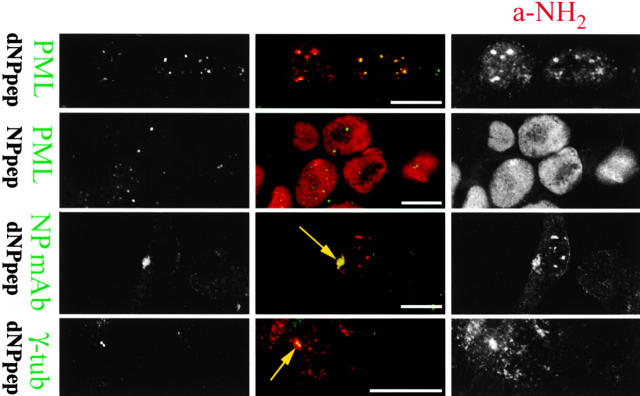
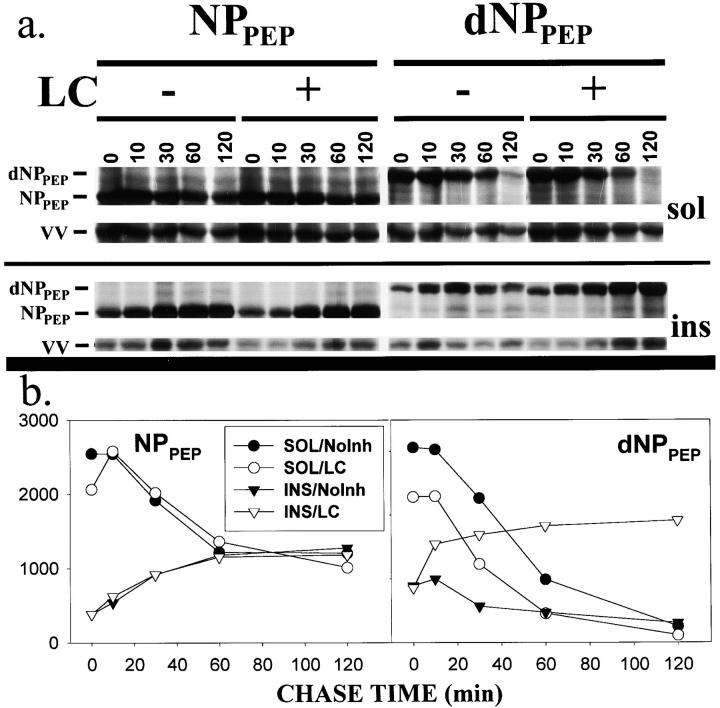
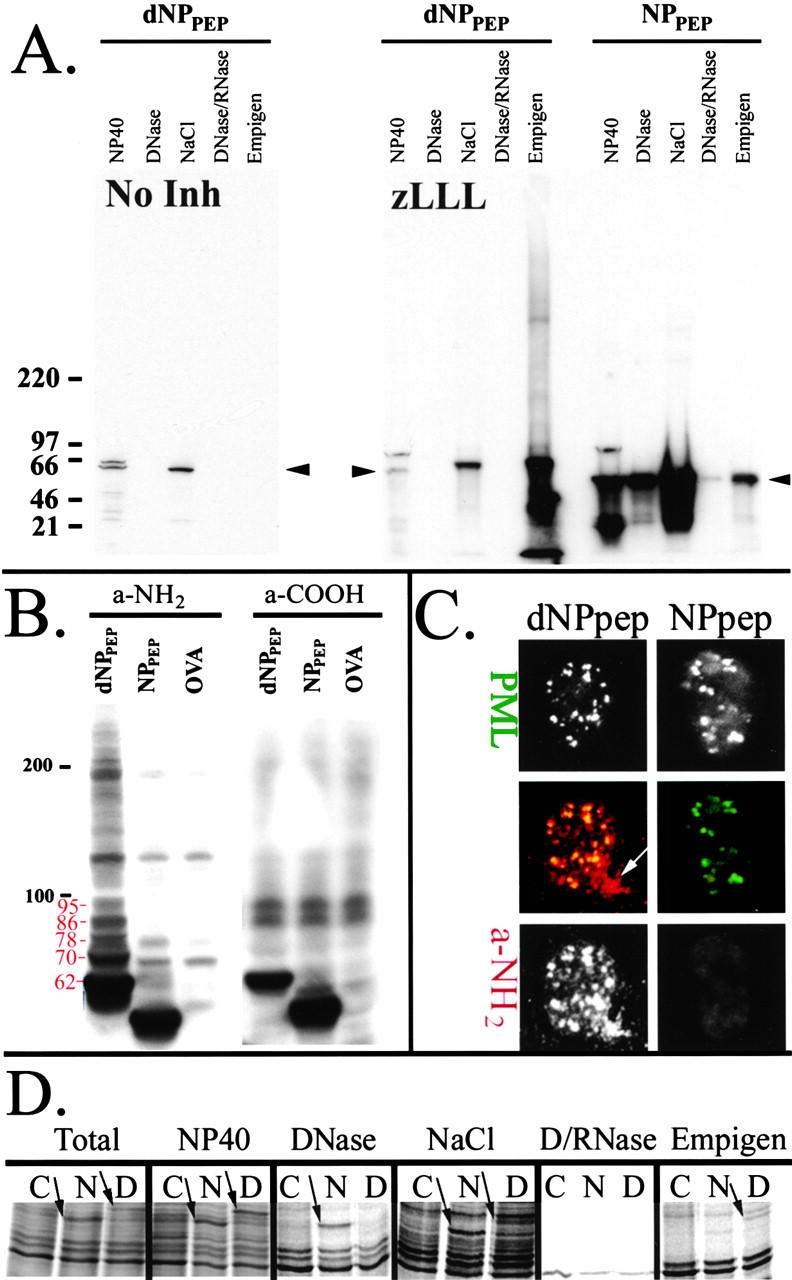
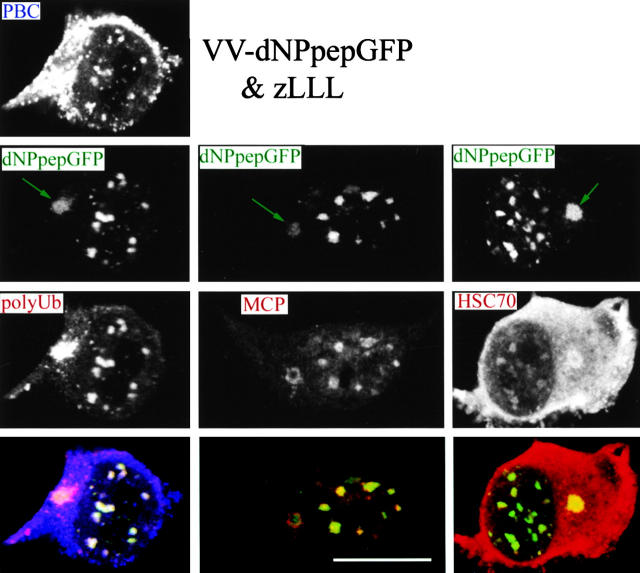
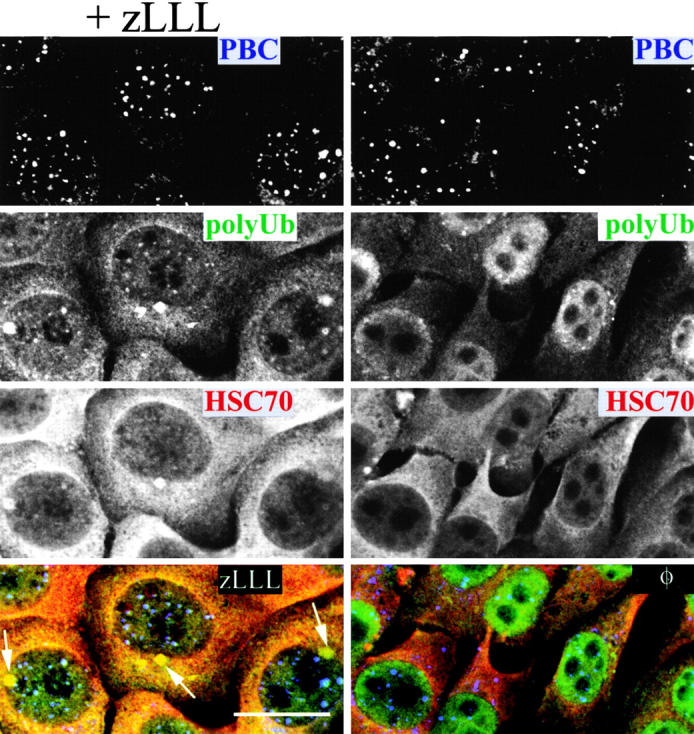
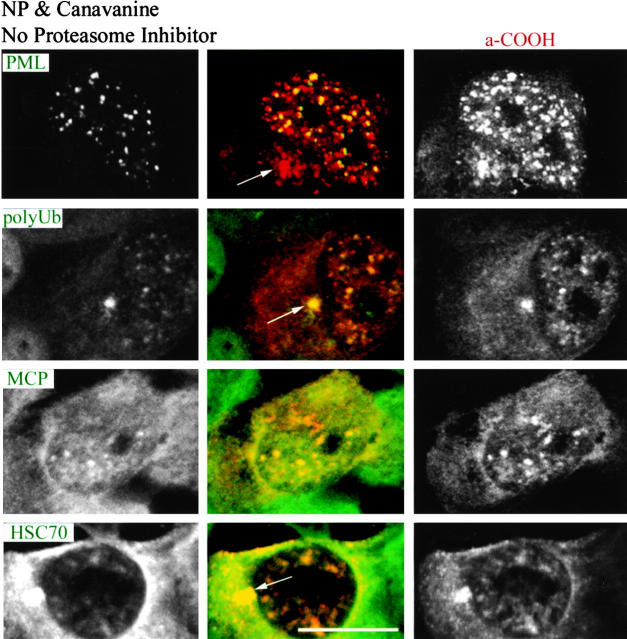
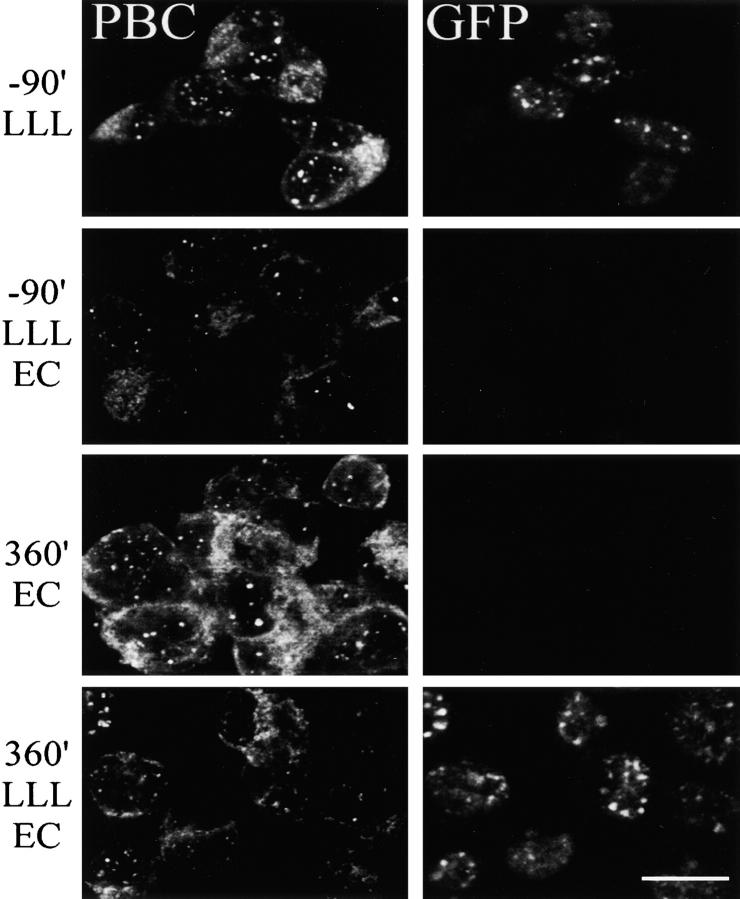
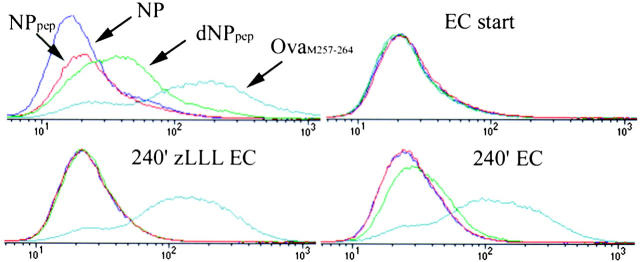
To better understand proteasomal degradation of nuclear proteins and viral antigens we studied mutated forms of influenza virus nucleoprotein (NP) that misfold and are rapidly degraded by proteasomes. In the presence of proteasome inhibitors, mutated NP (dNP) accumulates in highly insoluble ubiquitinated and nonubiquitinated species in nuclear substructures known as promyelocytic leukemia oncogenic domains (PODs) and the microtubule organizing center (MTOC). Immunofluorescence revealed that dNP recruits proteasomes and a selective assortment of molecular chaperones to both locales, and that a similar (though less dramatic) effect is induced by proteasome inhibitors in the absence of dNP expression. Biochemical evidence is consistent with the idea that dNP is delivered to PODs/MTOC in the absence of proteasome inhibitors. Restoring proteasome activity while blocking protein synthesis results in disappearance of dNP from PODs and the MTOC and the generation of a major histocompatibility complex class I-bound peptide derived from dNP but not NP. These findings demonstrate that PODs and the MTOC serve as sites of proteasomal degradation of misfolded dNP and probably cellular proteins as well, and imply that antigenic peptides are generated at one or both of these sites.
Figures
References
-
- Antón L.C., Snyder H.L., Bennink J.R., Vinitsky A., Orlowski M., Porgador A., Yewdell J.W. Dissociation of proteasomal degradation of biosynthesized viral proteins from generation of MHC class I-associated antigenic peptides. J. Immunol. 1998;160:4859–4868. - PubMed
-
- Bercovich B., Stancovski I., Mayer A., Blumenfeld N., Laszlo A., Schwartz A.L., Ciechanover A. Ubiquitin-dependent degradation of certain protein substrates in vitro requires the molecular chaperone Hsc70. J. Biol. Chem. 1997;272:9002–9010. - PubMed
-
- Bogyo M., Gaczynska M., Ploegh H.L. Proteasome inhibitors and antigen presentation. Biopolymers. 1997;43:269–280. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
