

The stoned proteins regulate synaptic vesicle recycling in the presynaptic terminal
- PMID: 10407025
- PMCID: PMC6783101
- DOI: 10.1523/JNEUROSCI.19-14-05847.1999
The stoned proteins regulate synaptic vesicle recycling in the presynaptic terminal
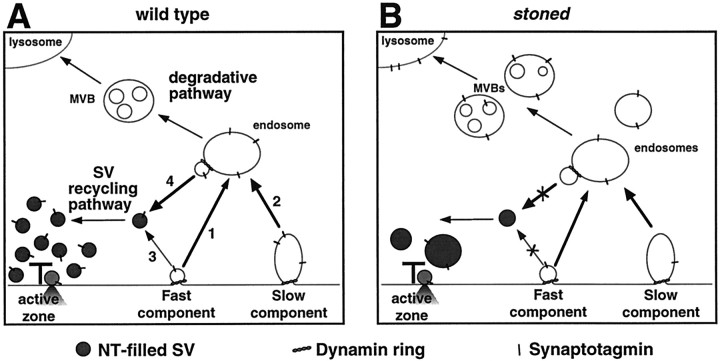
Abstract
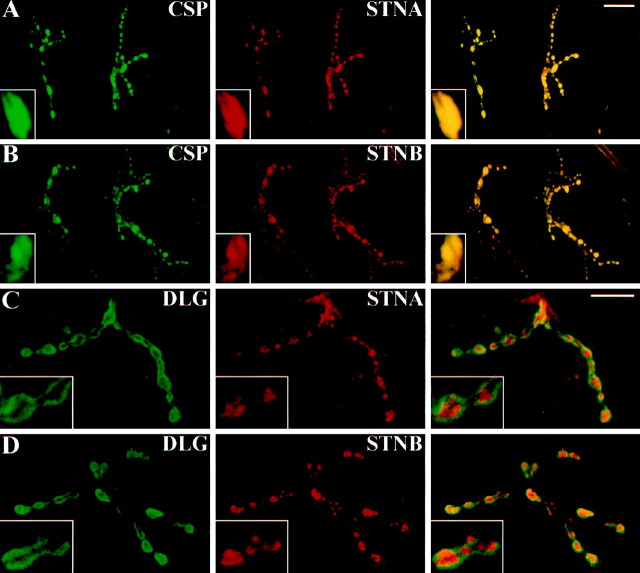
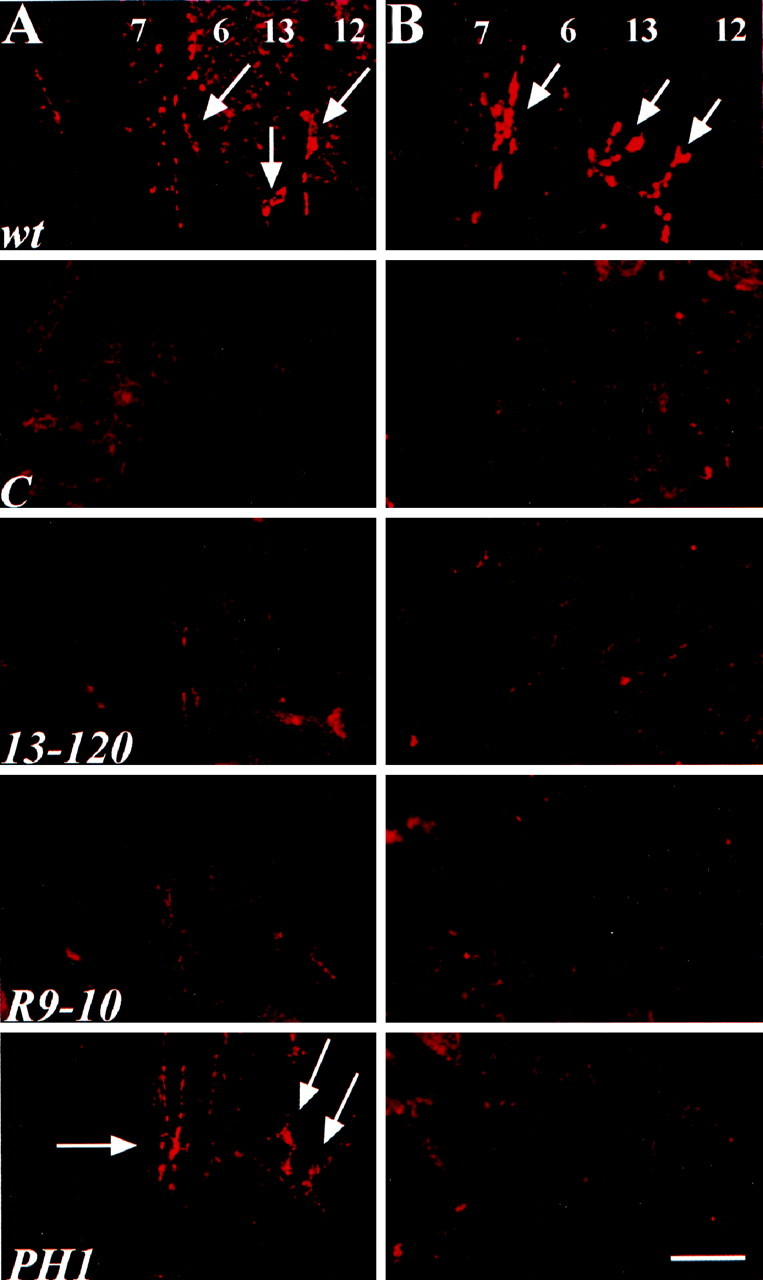
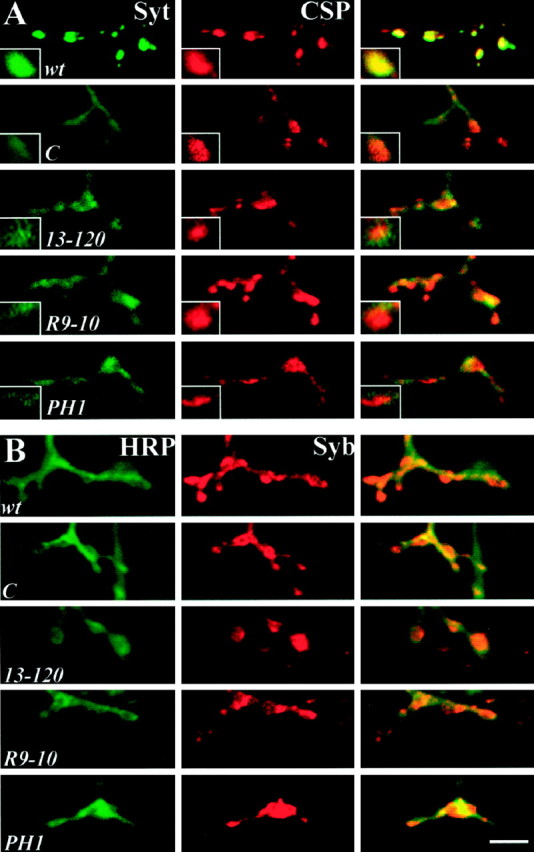
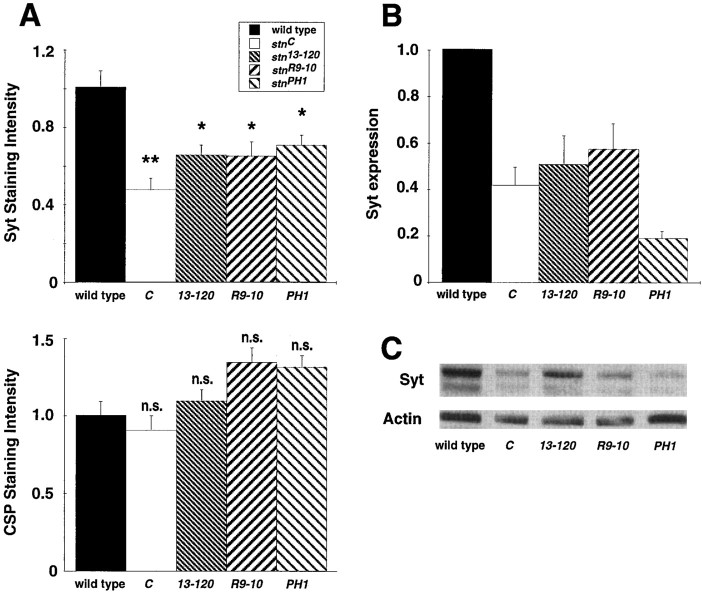
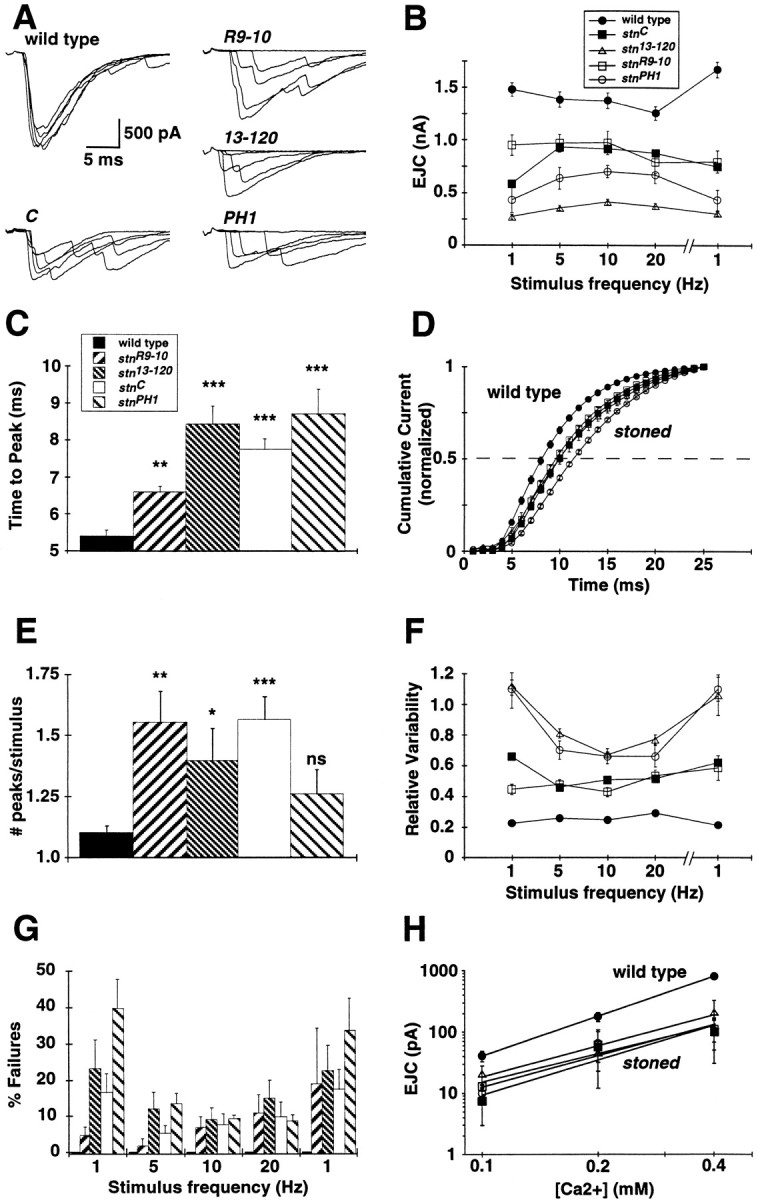
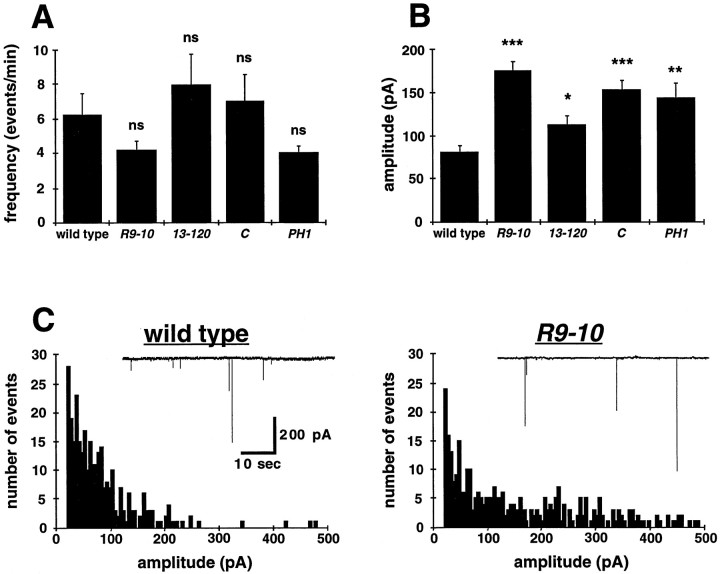
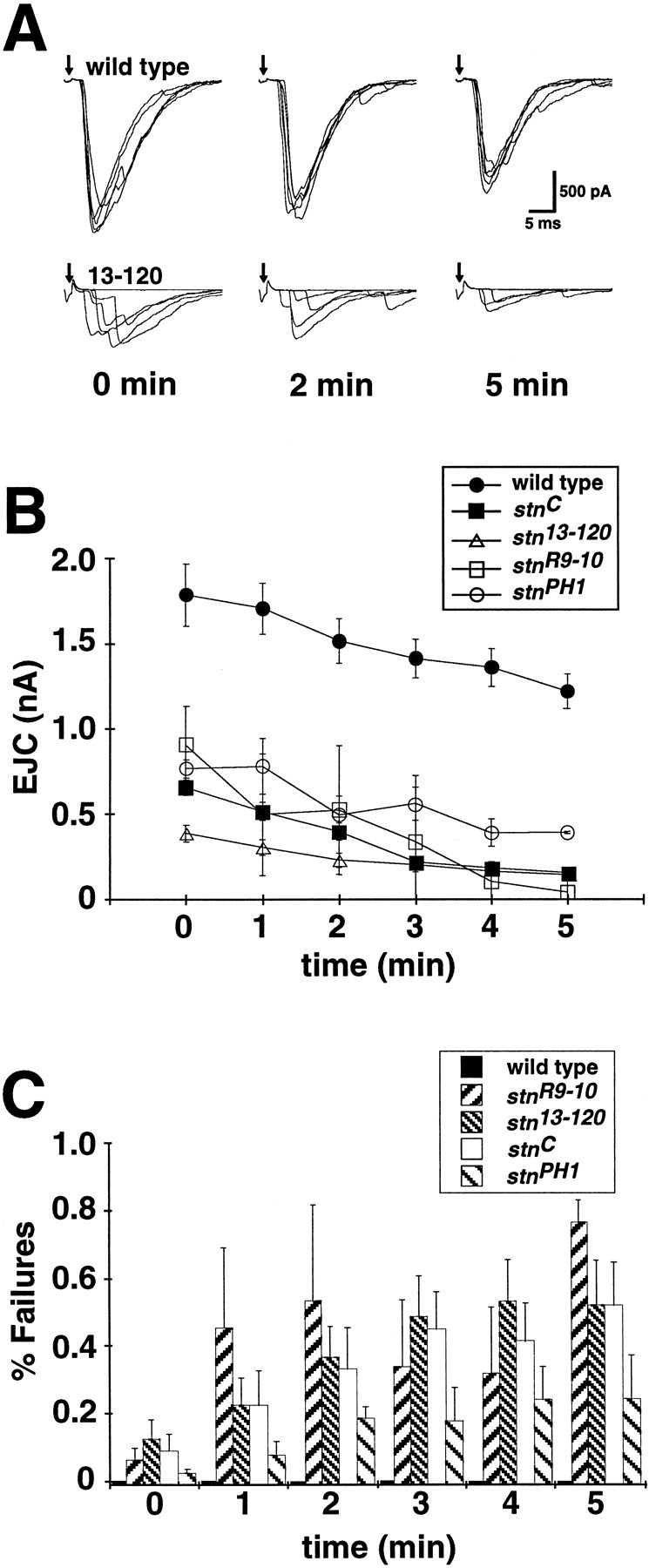
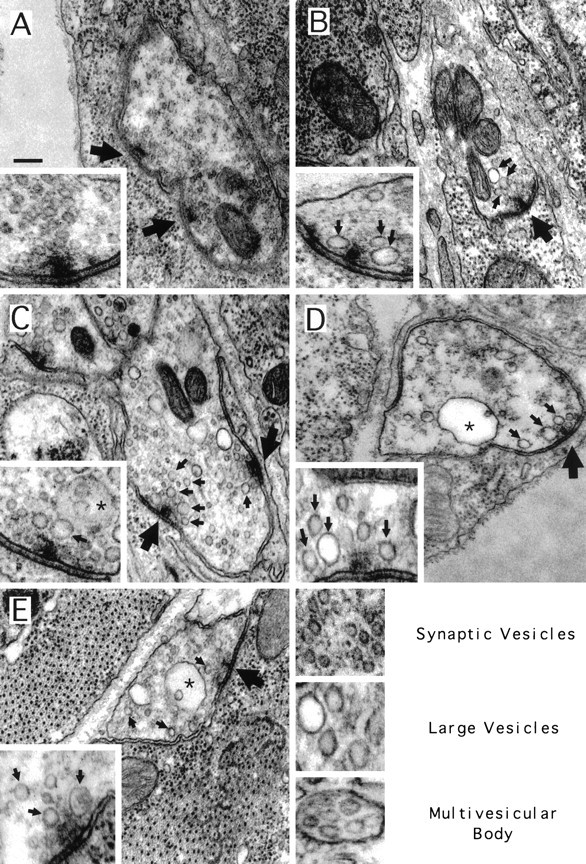
The Drosophila stoned locus was identified 25 years ago on the basis of stress-sensitive behavioral mutants (Grigliatti et al., 1973). The locus is dicistronic and encodes two distinct proteins, stoned A and stoned B, which are expressed specifically in presynaptic terminals at central and peripheral synapses. Several stoned mutant alleles cause embryonic lethality, suggesting that these proteins are essential for synaptic function. Physiological analyses at the stoned synapse reveal severe neurotransmission defects, including reduced and asynchronous neurotransmitter release and rapid fatigue after repetitive stimulation. At the EM level, stoned synapses show a depletion of synaptic vesicles and a concomitant increase in membrane-recycling intermediates. Mutant terminals also display a specific mislocalization of the synaptic vesicle protein synaptotagmin. These results suggest that the stoned proteins are essential for the recycling of synaptic vesicle membrane and are required for the proper sorting of synaptotagmin during endocytosis.
Figures
References
-
- Atwood HL, Govind CK, Wu CF. Differential ultrastructure of synaptic terminals on ventral longitudinal abdominal muscles in Drosophila larvae. J Neurobiol. 1993;24:1008–1024. - PubMed
-
- Bennett MK, Scheller RH. A molecular description of synaptic vesicle membrane trafficking. Annu Rev Biochem. 1994;63:63–100. - PubMed
-
- Betz WJ, Angleson JK. The synaptic vesicle cycle. Annu Rev Physiol. 1998;60:347–363. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases