

Impairment of AMPA receptor function in cerebellar granule cells of ataxic mutant mouse stargazer
- PMID: 10407040
- PMCID: PMC6783074
- DOI: 10.1523/JNEUROSCI.19-14-06027.1999
Impairment of AMPA receptor function in cerebellar granule cells of ataxic mutant mouse stargazer
Abstract
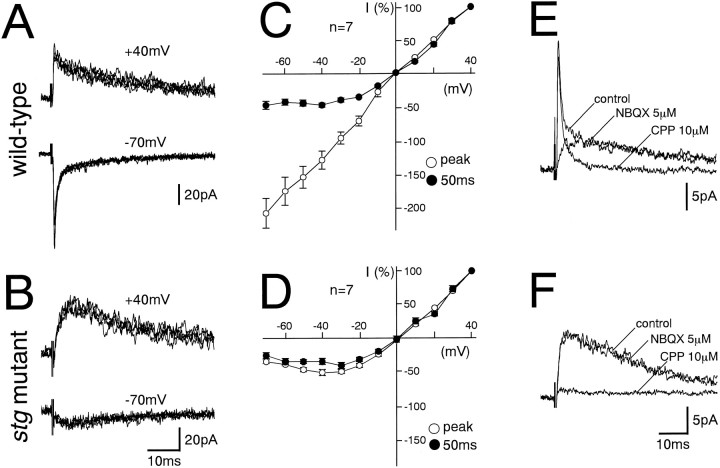
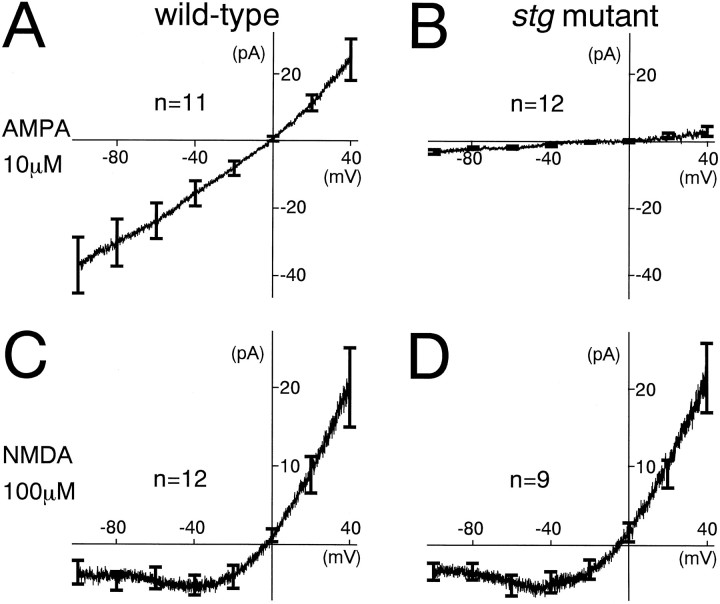
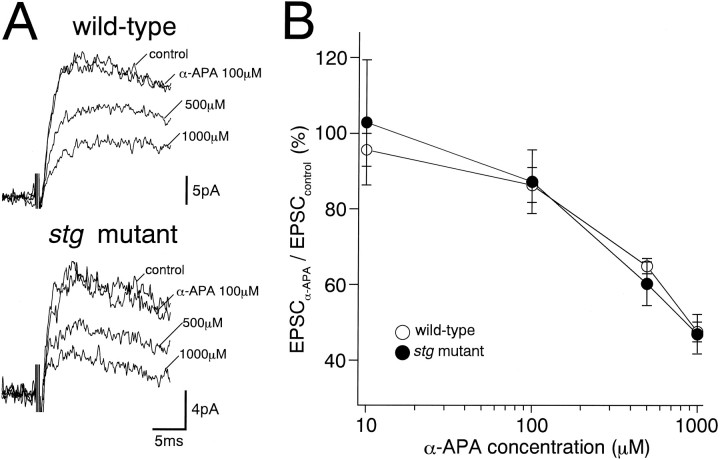
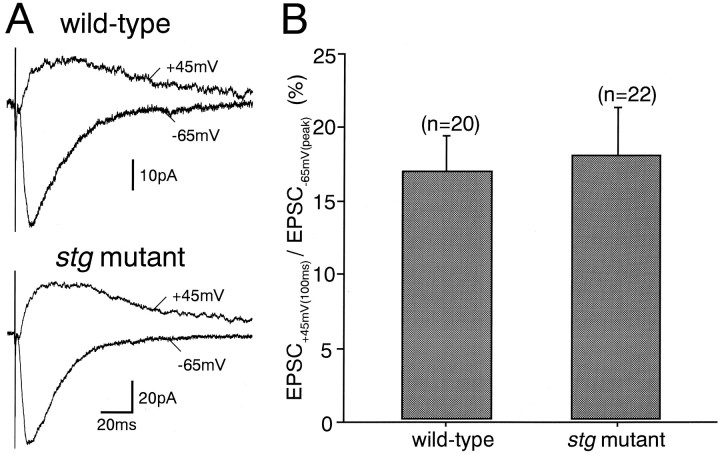
The spontaneous recessive mutant mouse stargazer (stg) begins to show ataxia around postnatal day 14 and display a severe impairment in the acquisition of classical eyeblink conditioning in adulthood. These abnormalities have been attributed to the specific reduction in brain-derived neurotrophic factor (BDNF) and the subsequent defect in TrkB receptor signaling in cerebellar granule cells (GCs). In the stg mutant cerebellum, we found that EPSCs at mossy fiber (MF) to GC synapses are devoid of the fast component mediated by AMPA-type glutamate receptors despite the normal slow component mediated by NMDA receptors. The sensitivity of stg mutant GCs to exogenously applied AMPA was greatly reduced, whereas that to NMDA was unchanged. Glutamate release from MF terminals during synaptic transmission to GCs appeared normal. By contrast, AMPA receptor-mediated EPSCs were normal in CA1 pyramidal cells of the stg mutant hippocampus. Thus, postsynaptic AMPA receptor function was selectively impaired in stg mutant GCs, although the transcription of four AMPA receptor subunit genes in the stg GC was comparable to the wild-type GC. We also examined the cerebellum of BDNF knockout mice and found that their MF-GC synapses had a normal AMPA receptor-mediated EPSC component. Thus, the impaired AMPA receptor function in the stg mutant GC is not likely to result from the reduced BDNF-TrkB signaling. These results suggest that the defect in MF to GC synaptic transmission is a major factor that causes the cerebellar dysfunction in the stg mutant mouse.
Figures
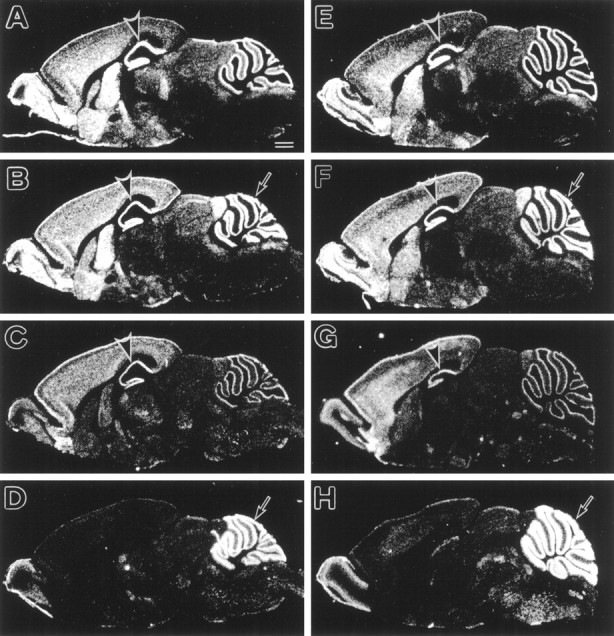
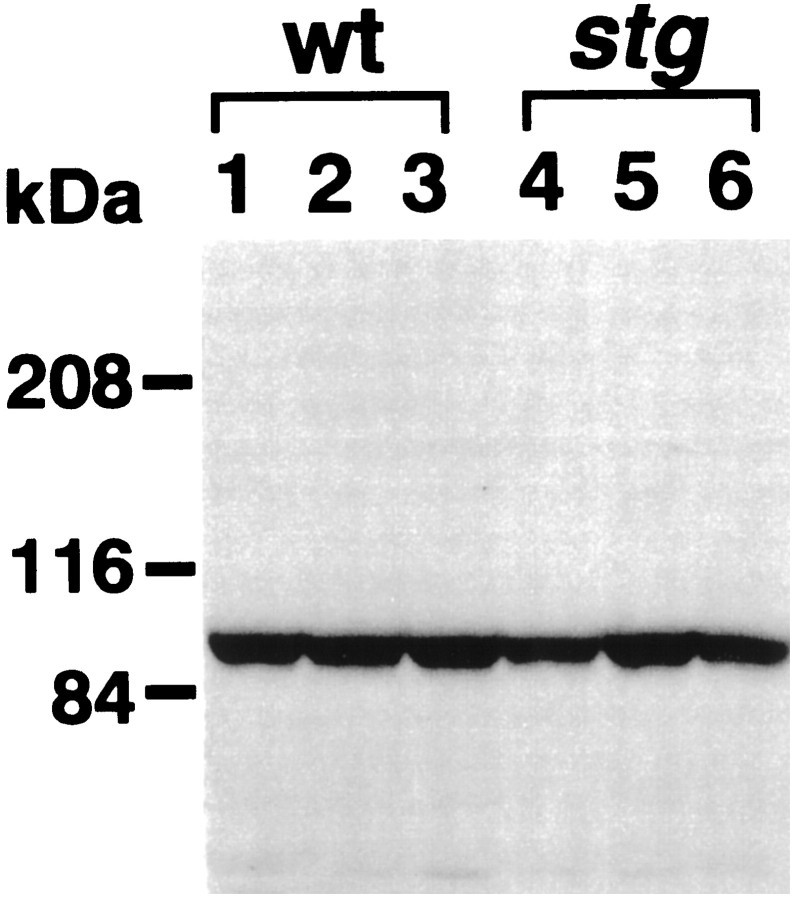
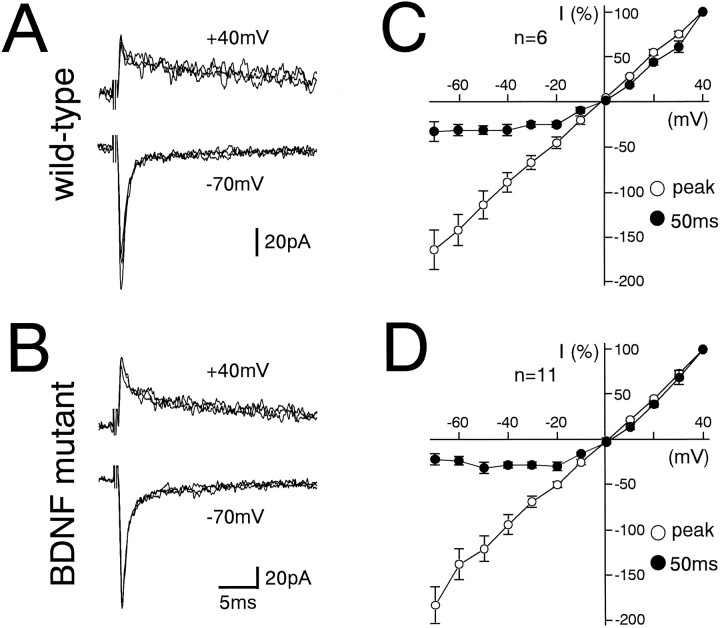
References
-
- Aiba A, Kano M, Chen C, Stanto ME, Fox GD, Herrup K, Zwingman TA, Tonegawa S. Deficient cerebellar long-term depression and impaired motor learning in mGluR1 mutant mice. Cell. 1994;79:377–388. - PubMed
-
- Boulter J, Hollmann M, O’Shea-Greenfield A, Hartley M, Deneris E, Maron C, Heinemann S. Molecular cloning and functional expression of glutamate receptor subunit genes. Science. 1990;249:1033–1037. - PubMed
-
- D’Angelo E, Rossi P, Taglietti V. Voltage-dependent kinetics of N-methyl-d-aspartate synaptic currents in rat cerebellar granule cells. Eur J Neurosci. 1994;6:640–645. - PubMed
-
- Di Pasquale E, Keegan KD, Noebels JL. Increased excitability and inward rectification in layer V cortical pyramidal neurons in epileptic mutant mouse stargazer. J Neurophysiol. 1997;77:621–631. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous