

Neural correlates of perceived brightness in the retina, lateral geniculate nucleus, and striate cortex
- PMID: 10407050
- PMCID: PMC6783067
- DOI: 10.1523/JNEUROSCI.19-14-06145.1999
Neural correlates of perceived brightness in the retina, lateral geniculate nucleus, and striate cortex
Abstract
Brightness changes can be induced in a static gray field by modulating the luminance of surrounding areas. We used this induction phenomenon to investigate the neural representation of perceived brightness. Extracellular recordings were made in striate cortex, the lateral geniculate nucleus (LGN), and the optic tract of anesthetized cats using stimuli that produced brightness induction. While a cell's receptive field (RF) was covered by uniform gray illumination, the luminance of rectangular flanking regions was modulated sinusoidally in time, inducing brightness changes in the RF. We looked for a correspondence between the modulation of a cell's response and stimulus conditions that did or did not produce perceptual changes in brightness. We found that the responses of retinal ganglion cell axons in the optic tract were never correlated with brightness. On the other hand, many neurons in striate cortex and a small fraction in the LGN responded in a phase-locked manner at the temporal frequency of the flank modulation, even though the flanks were 3-7 degrees beyond the edges of the RF. Only in striate cortex were cells found that had responses correlated with brightness under all stimulus conditions. These findings suggest that brightness information is explicitly represented in the responses of neurons in striate cortex as part of a neural representation of object surfaces.
Figures






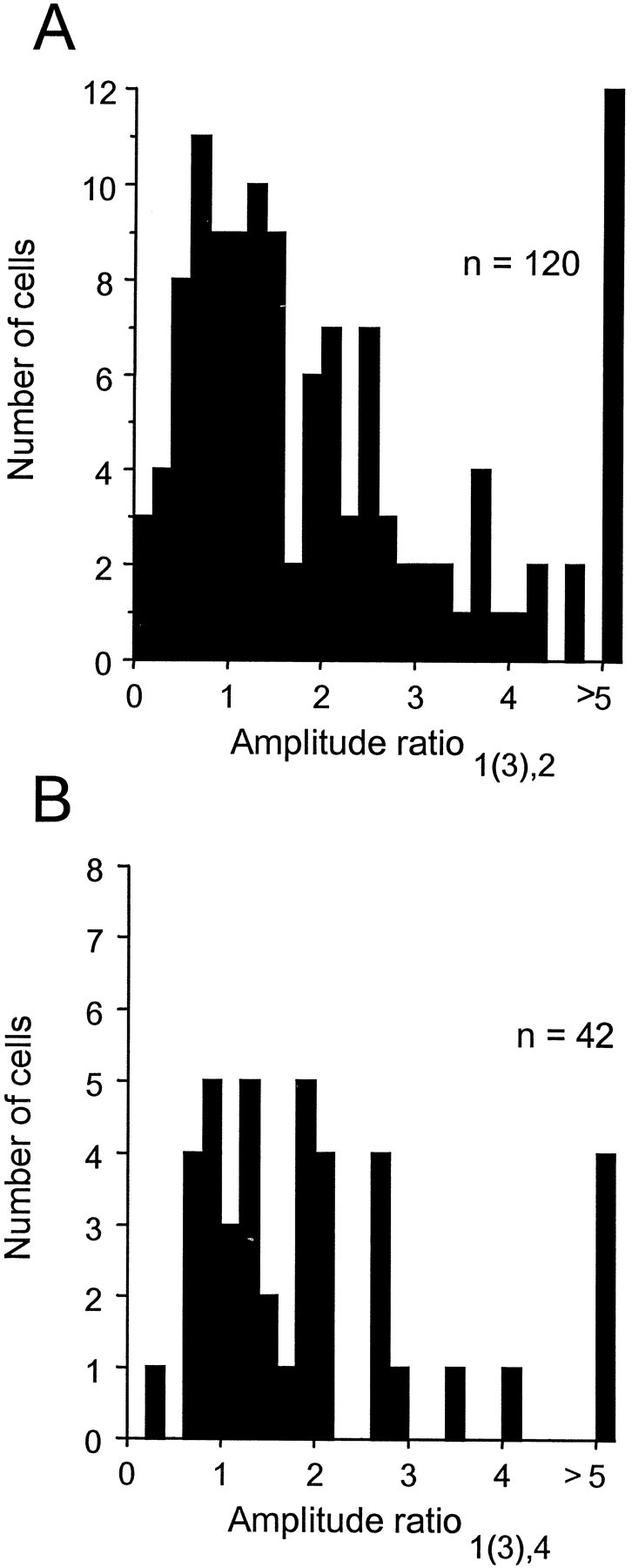
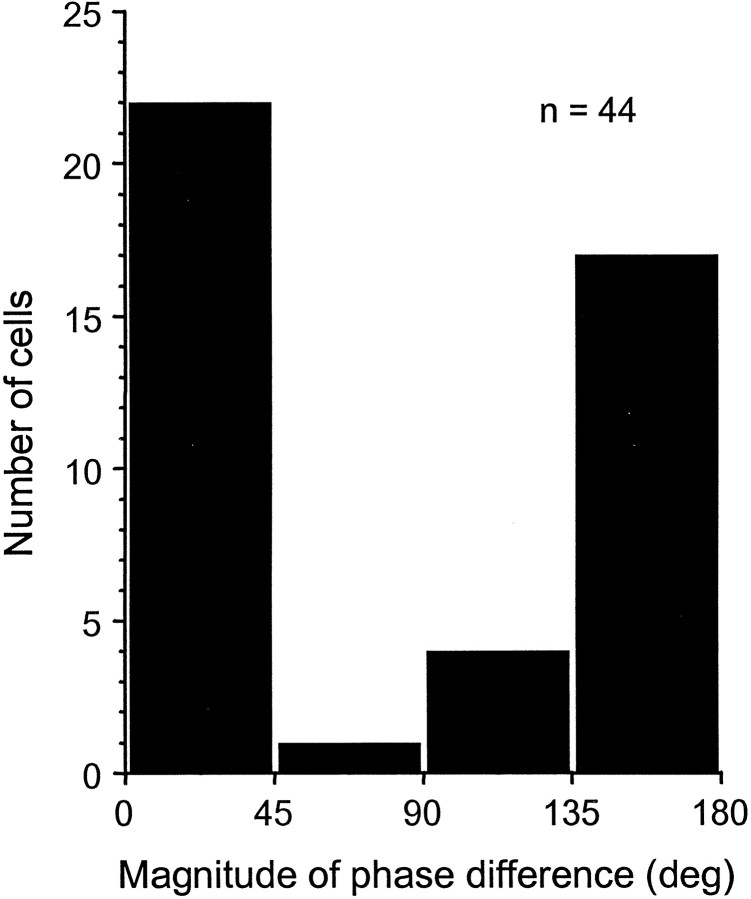




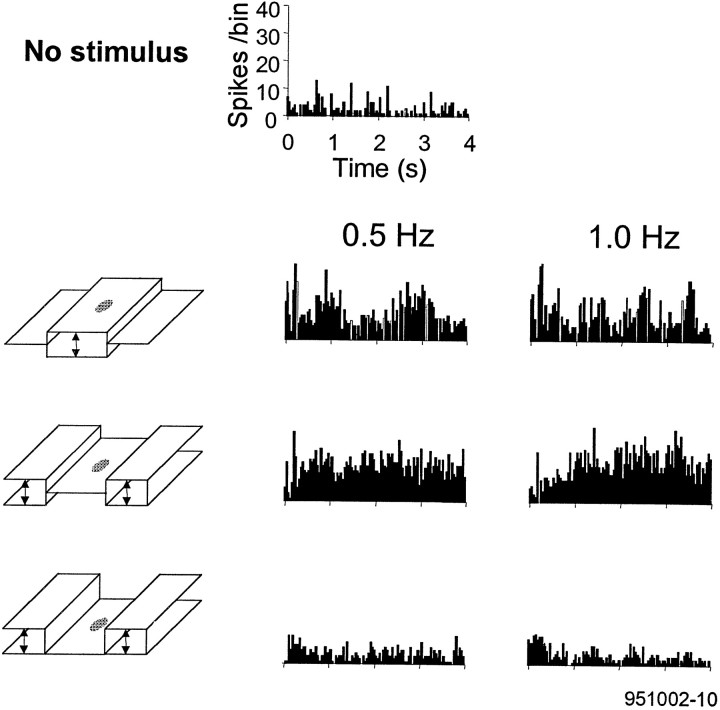
References
-
- Allman J, Miezin F, McGuinness E. Stimulus specific responses from beyond the classical receptive field: neurophysiological mechanisms for local-global comparisons in visual neurons. Annu Rev Neurosci. 1985;8:407–430. - PubMed
-
- Bravo M, Blake R, Morrison S. Cats see subjective contours. Vision Res. 1988;28:861–865. - PubMed
-
- DeAngelis GC, Robson JG, Ohzawa I, Freeman RD. Organization of suppression in receptive fields of neurons in cat visual cortex. J Neurophysiol. 1992;68:144–163. - PubMed
-
- DeValois RL, Pease PL. Contours and contrast: responses of monkey lateral geniculate nucleus cells to luminance and color figures. Science. 1971;171:694–696. - PubMed
-
- DeValois RL, Webster MA, De Valois KK, Lingelbach B. Temporal properties of brightness and color induction. Vision Res. 1986;26:887–897. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous