

Bradykinin B(2) receptor-mediated mitogen-activated protein kinase activation in COS-7 cells requires dual signaling via both protein kinase C pathway and epidermal growth factor receptor transactivation
- PMID: 10409720
- PMCID: PMC84372
- DOI: 10.1128/MCB.19.8.5289
Bradykinin B(2) receptor-mediated mitogen-activated protein kinase activation in COS-7 cells requires dual signaling via both protein kinase C pathway and epidermal growth factor receptor transactivation
Abstract
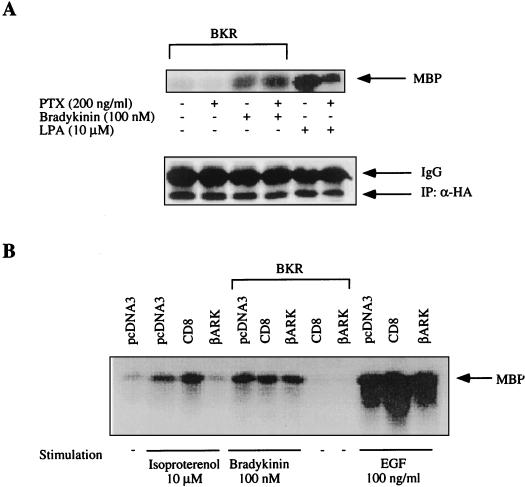
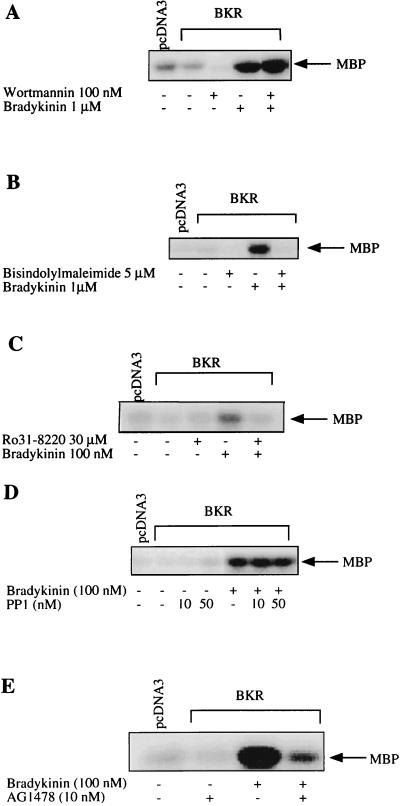
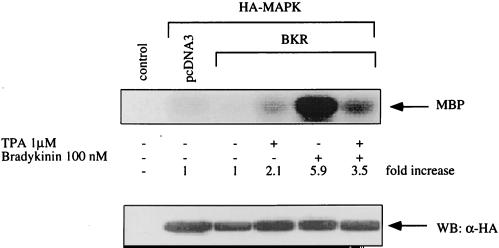
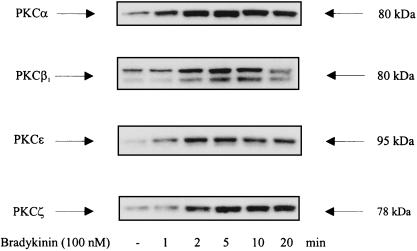
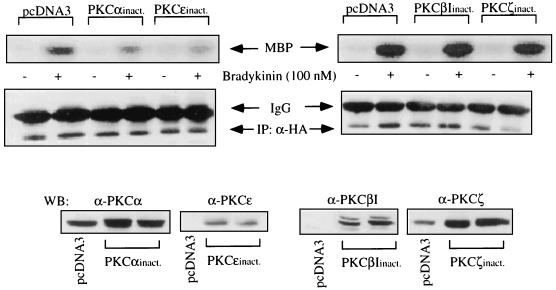
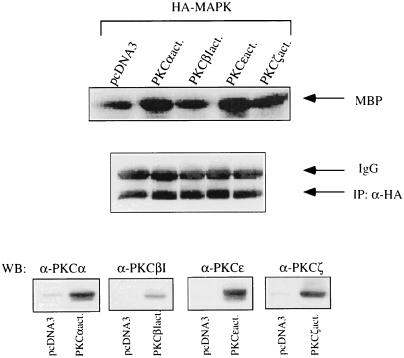
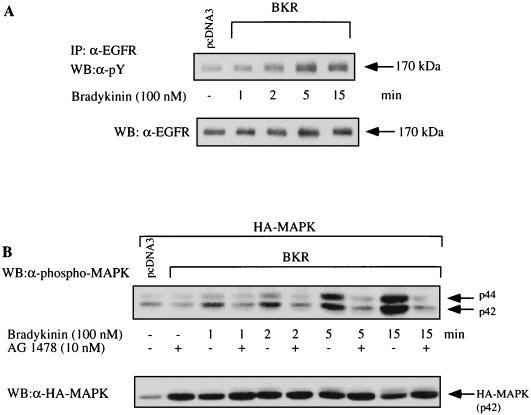
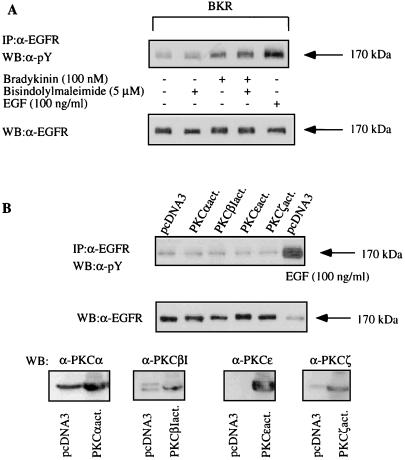
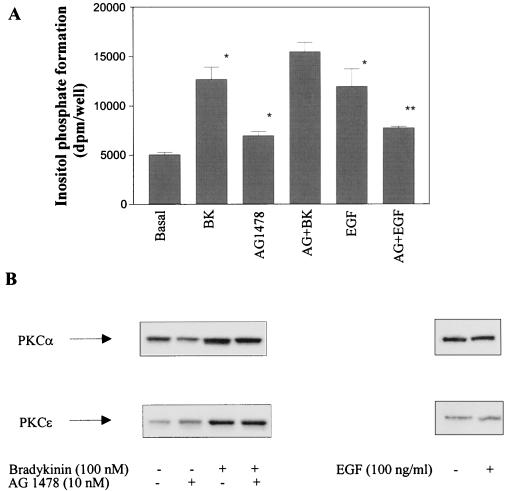
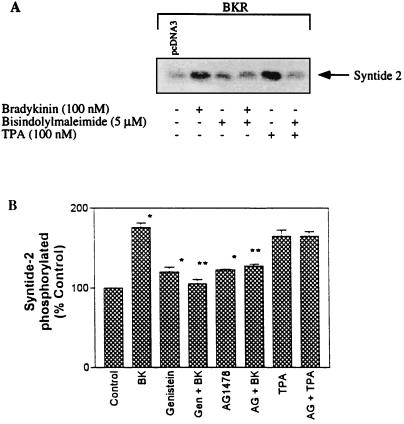
The signaling routes linking G-protein-coupled receptors to mitogen-activated protein kinase (MAPK) may involve tyrosine kinases, phosphoinositide 3-kinase gamma (PI3Kgamma), and protein kinase C (PKC). To characterize the mitogenic pathway of bradykinin (BK), COS-7 cells were transiently cotransfected with the human bradykinin B(2) receptor and hemagglutinin-tagged MAPK. We demonstrate that BK-induced activation of MAPK is mediated via the alpha subunits of a G(q/11) protein. Both activation of Raf-1 and activation of MAPK in response to BK were blocked by inhibitors of PKC as well as of the epidermal growth factor (EGF) receptor. Furthermore, in PKC-depleted COS-7 cells, the effect of BK on MAPK was clearly reduced. Inhibition of PI3-Kgamma or Src kinase failed to diminish MAPK activation by BK. BK-induced translocation and overexpression of PKC isoforms as well as coexpression of inactive or constitutively active mutants of different PKC isozymes provided evidence for a role of the diacylglycerol-sensitive PKCs alpha and epsilon in BK signaling toward MAPK. In addition to PKC activation, BK also induced tyrosine phosphorylation of EGF receptor (transactivation) in COS-7 cells. Inhibition of PKC did not alter BK-induced transactivation, and blockade of EGF receptor did not affect BK-stimulated phosphatidylinositol turnover or BK-induced PKC translocation, suggesting that PKC acts neither upstream nor downstream of the EGF receptor. Comparison of the kinetics of PKC activation and EGF receptor transactivation in response to BK also suggests simultaneous rather than consecutive signaling. We conclude that in COS-7 cells, BK activates MAPK via a permanent dual signaling pathway involving the independent activation of the PKC isoforms alpha and epsilon and transactivation of the EGF receptor. The two branches of this pathway may converge at the level of the Ras-Raf complex.
Figures
References
-
- Akiyama T, Ishida J, Nakagawa S, Ogawara H, Watanabe S, Itoh N, Shibuya M, Fukami Y. Genistein, a specific inhibitor of tyrosine-specific protein kinases. J Biol Chem. 1987;262:5592–5995. - PubMed
-
- Coutant K D, Corvaia N, Ryder N S. Bradykinin induces tyrosine phosphorylation of epidermal growth factor-receptor and focal adhesion proteins in human keratinocytes. Biochem Biophys Res Commun. 1995;210:774–780. - PubMed
-
- Crespo P, Xu N, Simonds W F, Gutkind J S. Ras-dependent activation of MAP kinase pathway mediated by G-protein beta gamma subunits. Nature. 1994;369:418–420. - PubMed
-
- Crespo P, Cachero T G, Xu N, Gutkind J S. Dual effect of beta-adrenergic receptors on mitogen-activated protein kinase. Evidence for a beta gamma-dependent activation and a G alpha s-cAMP-mediated inhibition. J Biol Chem. 1995;270:25259–25265. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous