

Quantitation of RNA polymerase II and its transcription factors in an HeLa cell: little soluble holoenzyme but significant amounts of polymerases attached to the nuclear substructure
- PMID: 10409729
- PMCID: PMC84381
- DOI: 10.1128/MCB.19.8.5383
Quantitation of RNA polymerase II and its transcription factors in an HeLa cell: little soluble holoenzyme but significant amounts of polymerases attached to the nuclear substructure
Abstract
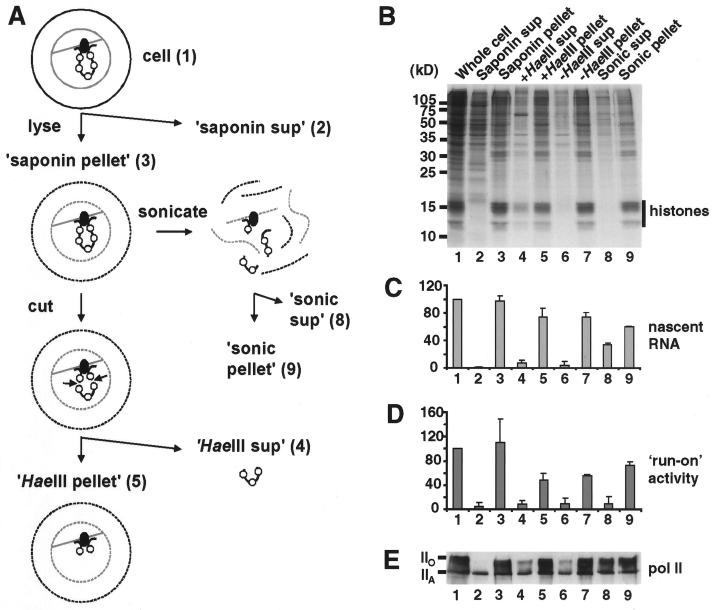
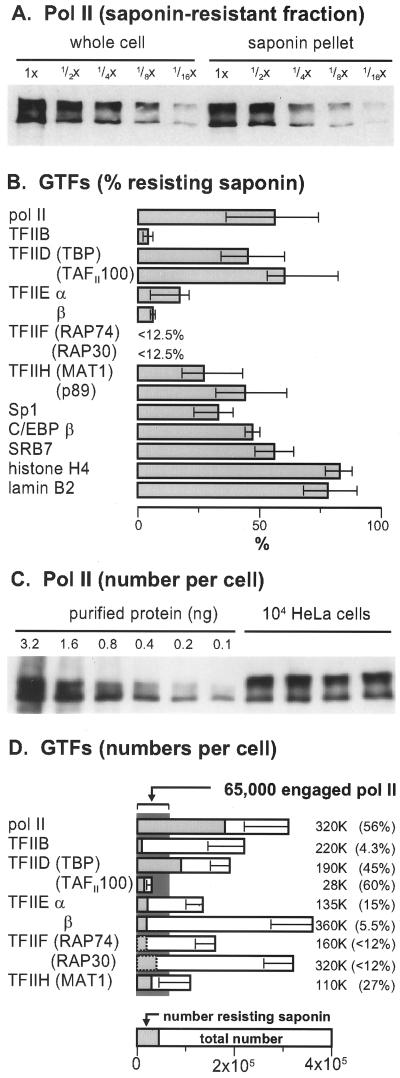
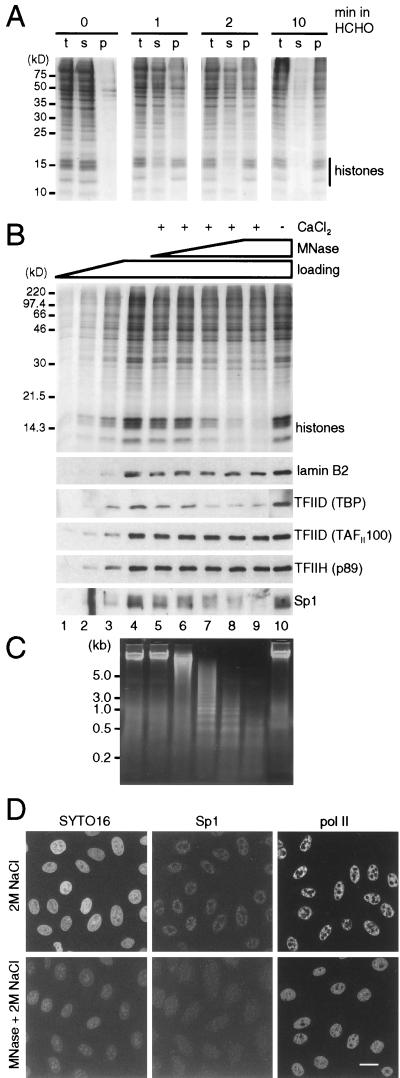
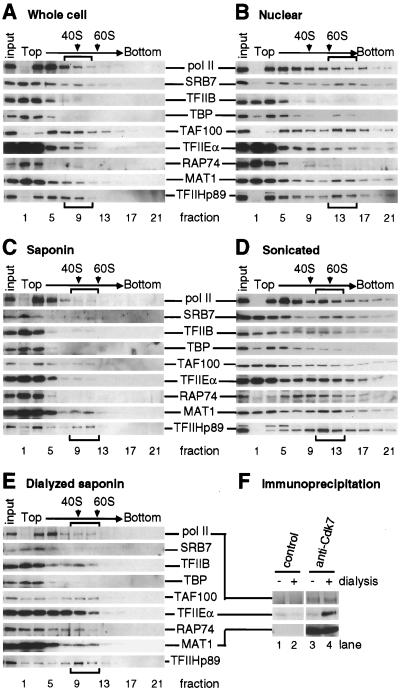
Various complexes that contain the core subunits of RNA polymerase II associated with different transcription factors have been isolated from eukaryotes; their precise molecular constitution depends on the purification procedure. We estimated the numbers of various components of such complexes in an HeLa cell by quantitative immunoblotting. The cells were lysed with saponin in a physiological buffer; approximately 140,000 unengaged polymerases (mainly of form IIA) were released. Only approximately 4,000 of these soluble molecules sedimented in glycerol gradients as holoenzyme-sized complexes. About 180,000 molecules of polymerases (approximately 110,000 molecules of form IIO) and 10,000 to 30,000 molecules of each of TFIIB, TFIIEalpha, TFIIEbeta, TFIIF-RAP74, TFIIF-RAP30, and TFIIH-MAT1 remained tightly associated with the nuclear substructure. Most proteins and run-on activity were retained when approximately 50% of the chromatin was detached with a nuclease, but approximately 45,000 molecules of bound TATA binding protein (TBP) were detached. Similar results were obtained after cross-linking living cells with formaldehyde. The results provide little support for the existence of a large pool of soluble holoenzyme; they are consistent with TBP-promoter complexes in nuclease-sensitive chromatin being assembled into preinitiation complexes attached to the underlying structure.
Figures


References
-
- Apone L M, Virbasius C M, Reese J C, Green M R. Yeast TAF(II)90 is required for cell-cycle progression through G2/M but not for general transcription activation. Genes Dev. 1996;10:2368–2380. - PubMed
-
- Barberis A, Pearlberg J, Simkovich N, Farrell S, Reinagel P, Bamdad C, Sigal G, Ptashne M. Contact with a component of the polymerase II holoenzyme suffices for gene activation. Cell. 1995;81:359–368. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials