

The cytosolic tail of class I MHC heavy chain is required for its dislocation by the human cytomegalovirus US2 and US11 gene products
- PMID: 10411907
- PMCID: PMC17548
- DOI: 10.1073/pnas.96.15.8516
The cytosolic tail of class I MHC heavy chain is required for its dislocation by the human cytomegalovirus US2 and US11 gene products
Abstract
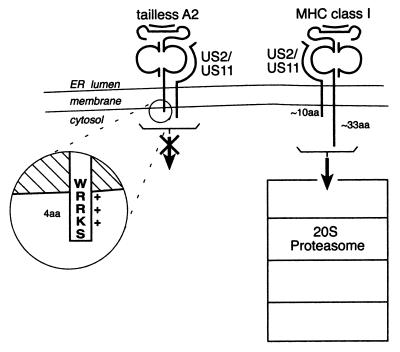
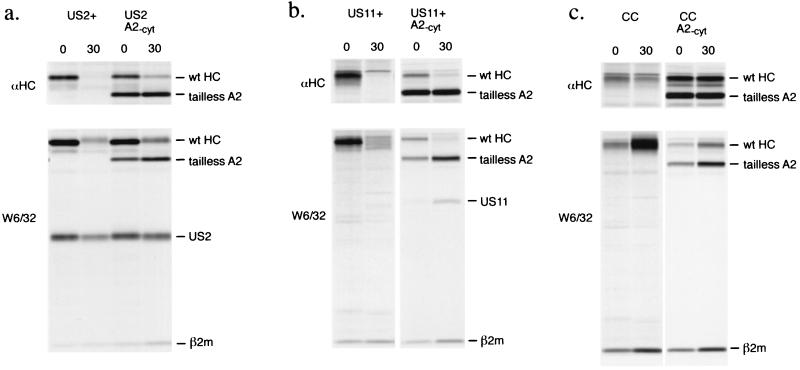
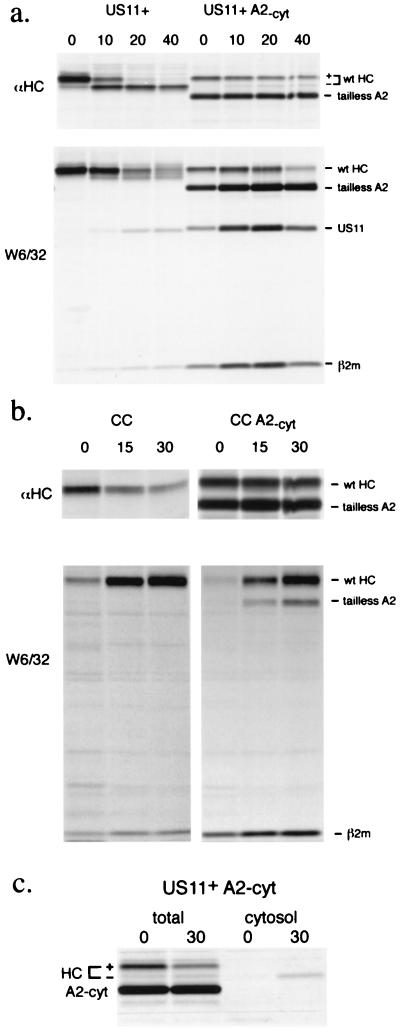
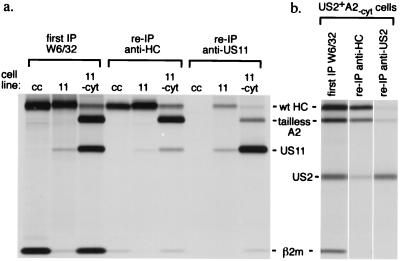
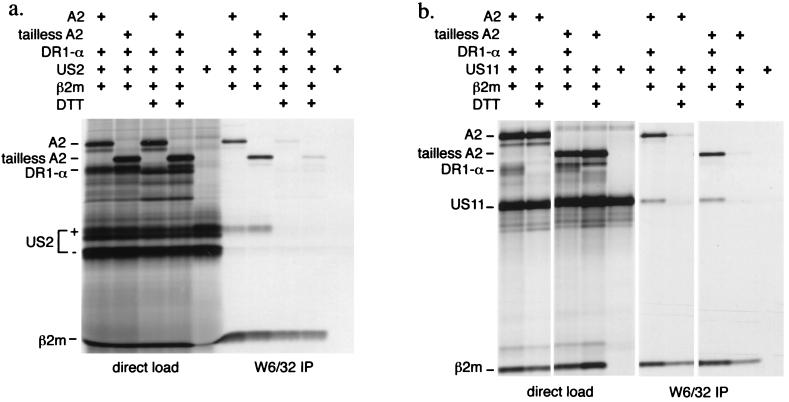
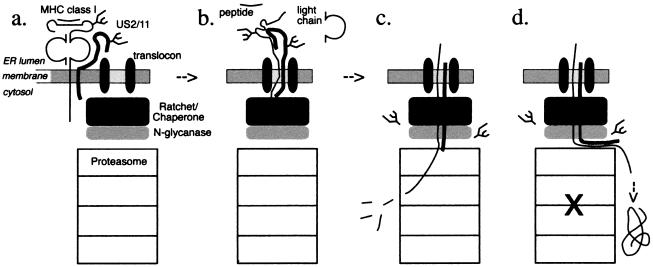
The US2 and US11 glycoproteins of human cytomegalovirus facilitate destruction of MHC class I heavy chains by proteasomal proteolysis through acceleration of endoplasmic reticulum-to-cytosol dislocation. Modification of the class I heavy chain was used to probe the structural requirements for this sequence of reactions. The cytosolic domain of the class I heavy chain is required for dislocation to the cytosol and for its subsequent destruction. However, interactions between US2 or US11 and the heavy chain are maintained in the absence of the class I cytosolic domain, as shown by chemical crosslinking in vivo and coprecipitation when translated in vitro. Thus, substrate recognition and accelerated destruction of the heavy chain, as facilitated by US2 or US11, are separable events.
Figures
Similar articles
-
Membrane-specific, host-derived factors are required for US2- and US11-mediated degradation of major histocompatibility complex class I molecules.J Biol Chem. 2002 Feb 1;277(5):3258-67. doi: 10.1074/jbc.M109765200. Epub 2001 Nov 20. J Biol Chem. 2002. PMID: 11717308
-
TRAM1 participates in human cytomegalovirus US2- and US11-mediated dislocation of an endoplasmic reticulum membrane glycoprotein.J Biol Chem. 2009 Feb 27;284(9):5905-14. doi: 10.1074/jbc.M807568200. Epub 2009 Jan 2. J Biol Chem. 2009. PMID: 19121997 Free PMC article.
-
Visualization of the ER-to-cytosol dislocation reaction of a type I membrane protein.EMBO J. 2002 Mar 1;21(5):1041-53. doi: 10.1093/emboj/21.5.1041. EMBO J. 2002. PMID: 11867532 Free PMC article.
-
The HCMV gene products US2 and US11 target MHC class I molecules for degradation in the cytosol.Curr Top Microbiol Immunol. 2002;269:37-55. doi: 10.1007/978-3-642-59421-2_3. Curr Top Microbiol Immunol. 2002. PMID: 12224515 Review.
-
Identifying the ERAD ubiquitin E3 ligases for viral and cellular targeting of MHC class I.Mol Immunol. 2015 Dec;68(2 Pt A):106-11. doi: 10.1016/j.molimm.2015.07.005. Epub 2015 Jul 22. Mol Immunol. 2015. PMID: 26210183 Free PMC article. Review.
Cited by
-
Regulation of ubiquitin-proteasome system mediated degradation by cytosolic stress.Mol Biol Cell. 2007 Nov;18(11):4279-91. doi: 10.1091/mbc.e07-05-0487. Epub 2007 Aug 15. Mol Biol Cell. 2007. PMID: 17699585 Free PMC article.
-
Human cytomegalovirus protein US11 provokes an unfolded protein response that may facilitate the degradation of class I major histocompatibility complex products.J Virol. 2005 Mar;79(5):2768-79. doi: 10.1128/JVI.79.5.2768-2779.2005. J Virol. 2005. PMID: 15708995 Free PMC article.
-
The VCP/p97 and YOD1 Proteins Have Different Substrate-dependent Activities in Endoplasmic Reticulum-associated Degradation (ERAD).J Biol Chem. 2015 Nov 20;290(47):28175-28188. doi: 10.1074/jbc.M115.656660. Epub 2015 Oct 13. J Biol Chem. 2015. PMID: 26463207 Free PMC article.
-
Dislocation of a type I membrane protein requires interactions between membrane-spanning segments within the lipid bilayer.Mol Biol Cell. 2003 Sep;14(9):3690-8. doi: 10.1091/mbc.e03-03-0192. Epub 2003 Jun 13. Mol Biol Cell. 2003. PMID: 12972557 Free PMC article.
-
Binding of human cytomegalovirus US2 to major histocompatibility complex class I and II proteins is not sufficient for their degradation.J Virol. 2002 Aug;76(16):8265-75. doi: 10.1128/jvi.76.16.8265-8275.2002. J Virol. 2002. PMID: 12134032 Free PMC article.
References
-
- Ploegh H L. Science. 1998;280:248–253. - PubMed
-
- Wiertz E J, Jones T R, Sun L, Bogyo M, Geuze H J, Ploegh H L. Cell. 1996;84:769–779. - PubMed
-
- Wiertz E J, Tortorella D, Bogyo M, Yu J, Mothes W, Jones T R, Rapoport T A, Ploegh H L. Nature (London) 1996;384:432–438. - PubMed
-
- Hiller M M, Finger A, Schweiger M, Wolf D H. Science. 1996;273:1725–1728. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
