

Dipeptidyl peptidase I is required for the processing and activation of granzymes A and B in vivo
- PMID: 10411926
- PMCID: PMC17567
- DOI: 10.1073/pnas.96.15.8627
Dipeptidyl peptidase I is required for the processing and activation of granzymes A and B in vivo
Abstract
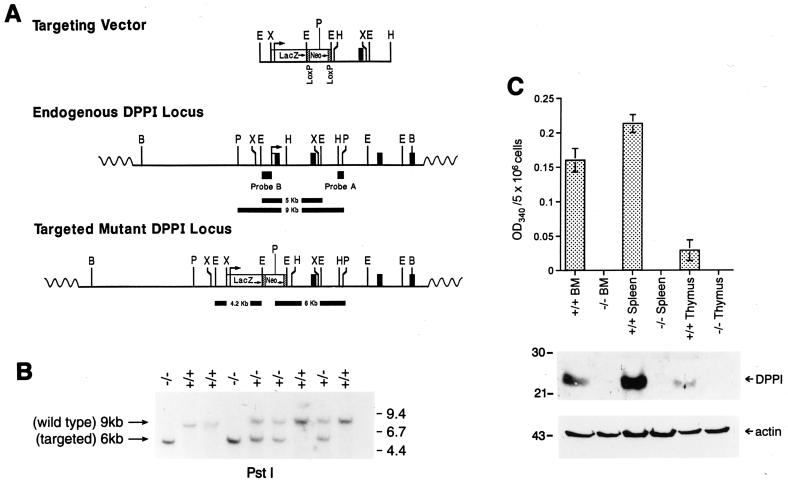
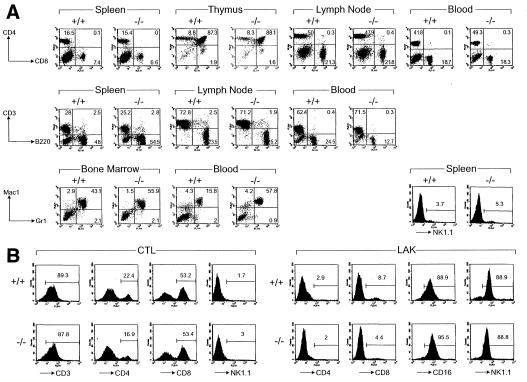
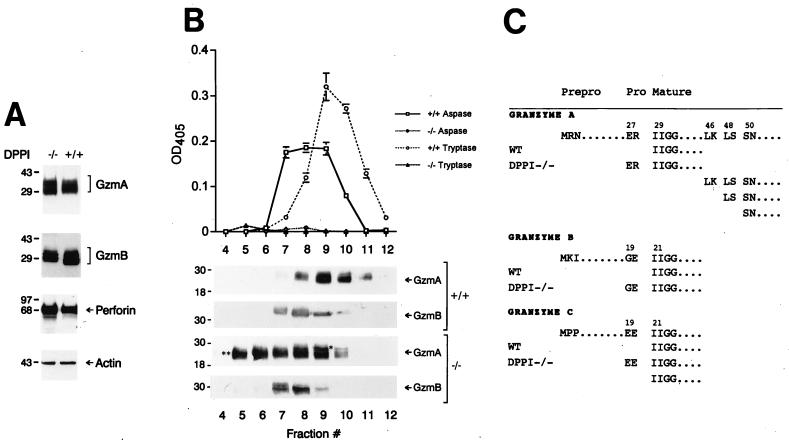
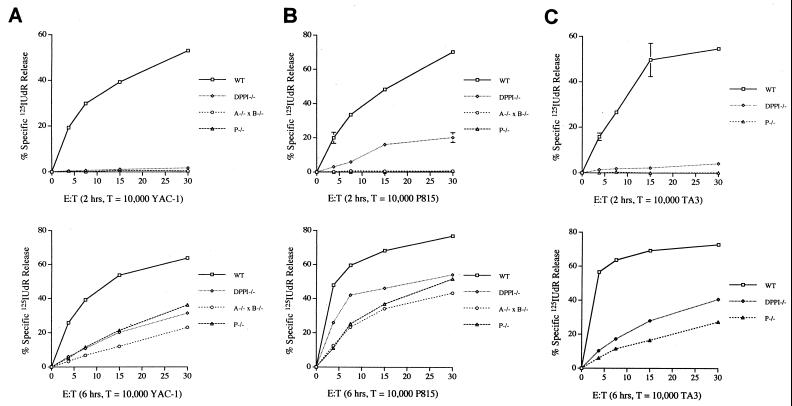
Dipeptidyl peptidase I (DPPI) is a lysosomal cysteine protease that has been implicated in the processing of granzymes, which are neutral serine proteases exclusively expressed in the granules of activated cytotoxic lymphocytes. In this report, we show that cytotoxic lymphocytes derived from DPPI-/- mice contain normal amounts of granzymes A and B, but these molecules retain their prodipeptide domains and are inactive. Cytotoxic assays with DPPI-/- effector cells reveal severe defects in the induction of target cell apoptosis (as measured by [(125)I]UdR release) at both early and late time points; this defect is comparable to that detected in perforin-/- or granzyme A-/- x B-/- cytotoxic lymphocytes. DPPI therefore plays an essential role in the in vivo processing and activation of granzymes A and B, which are required for cytotoxic lymphocyte granule-mediated apoptosis.
Figures
Comment in
-
How to induce involuntary suicide: the need for dipeptidyl peptidase I.Proc Natl Acad Sci U S A. 1999 Jul 20;96(15):8312-4. doi: 10.1073/pnas.96.15.8312. Proc Natl Acad Sci U S A. 1999. PMID: 10411865 Free PMC article. No abstract available.
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
