

The violaxanthin cycle protects plants from photooxidative damage by more than one mechanism
- PMID: 10411949
- PMCID: PMC17590
- DOI: 10.1073/pnas.96.15.8762
The violaxanthin cycle protects plants from photooxidative damage by more than one mechanism
Abstract
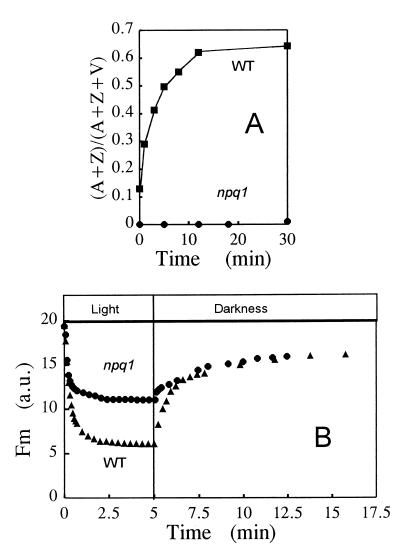
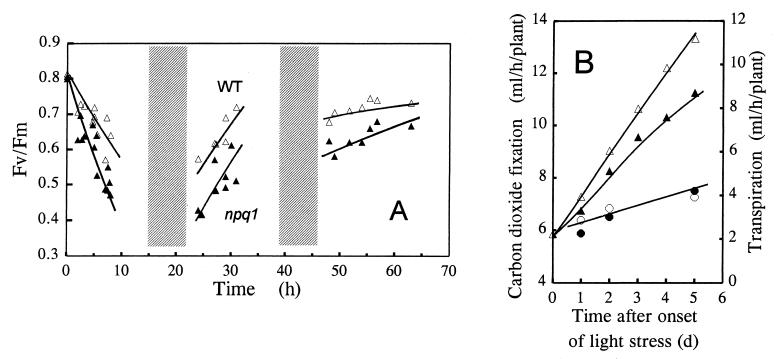
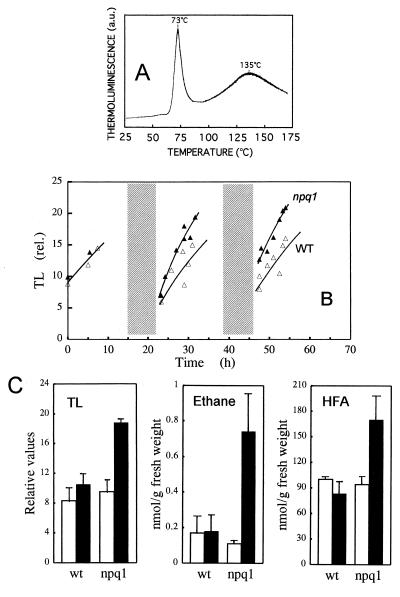
When light energy absorbed by plants becomes excessive relative to the capacity of photosynthesis, the xanthophyll violaxanthin is reversibly deepoxidized to zeaxanthin (violaxanthin cycle). The protective function of this phenomenon was investigated in a mutant of Arabidopsis thaliana, npq1, that has no functional violaxanthin deepoxidase. Two major consequences of the npq1 mutation are the absence of zeaxanthin formation in strong light and the partial inhibition of the quenching of singlet excited chlorophylls in the photosystem II light-harvesting complexes. Prolonged exposure of whole plants to bright light resulted in a limited photoinhibition of photosystem II in both npq1 and wild-type leaves, although CO(2) fixation and the linear electron transport in npq1 plants were reduced substantially. Lipid peroxidation was more pronounced in npq1 compared with the wild type, as measured by chlorophyll thermoluminescence, ethane production, and the total hydroperoxy fatty acids content. Lipid peroxidation was amplified markedly under chilling stress, and photooxidative damage ultimately resulted in leaf bleaching and tissue necrosis in npq1. The npq4 mutant, which possesses a normal violaxanthin cycle but has a limited capacity of quenching singlet excited chlorophylls, was rather tolerant to lipid peroxidation. The double mutant, npq4 npq1, which differs from npq4 only by the absence of the violaxanthin cycle, exhibited an increased susceptibility to photooxidative damage, similar to that of npq1. Our results demonstrate that the violaxanthin cycle specifically protects thylakoid membrane lipids against photooxidation. Part of this protection involves a mechanism other than quenching of singlet excited chlorophylls.
Figures

References
-
- Asada K, Takahashi M. In: Photoinhibition. Kyle D J, Osmond C B, Arntzen C J, editors. Amsterdam: Elsevier; 1987. pp. 227–287.
-
- Niyogi, K. K. (1999) Annu. Rev. Plant Physiol. Plant Mol. Biol.50, in press. - PubMed
-
- Demmig-Adams B, Adams W W., III Annu Rev Plant Physiol Plant Mol Biol. 1992;43:599–626.
-
- Barber J, Andersson B. Trends Biochem Sci. 1992;17:61–66. - PubMed
-
- Prasil O, Adir N, Ohad I. In: The Photosystems: Structure, Function and Molecular Biology. Barber J, editor. Amsterdam: Elsevier; 1992. pp. 295–348.
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
