

Review
doi: 10.1128/JB.181.15.4443-4451.1999.
Iron acquisition and metabolism by mycobacteria
Affiliations
- PMID: 10419938
- PMCID: PMC103571
- DOI: 10.1128/JB.181.15.4443-4451.1999
Item in Clipboard
Review
Iron acquisition and metabolism by mycobacteria
J Bacteriol.
1999 Aug.
No abstract available
Figures
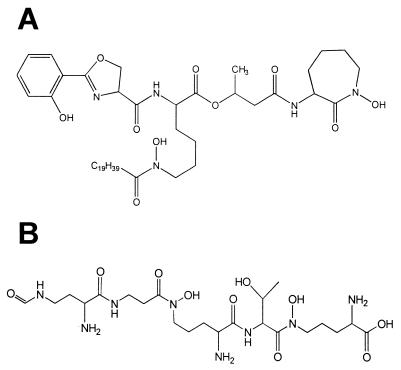
Siderophores of mycobacterial species. (A) Structure of the major MB of M. tuberculosis (MB T). (B) Structure of the major exochelin of M. smegmatis (exochelin MS). Peptidic siderophores such as exochelin MS have only been identified from fast-growing saprophytic strains, while MBs such as that shown in panel A have been found widely distributed among mycobacterial species.
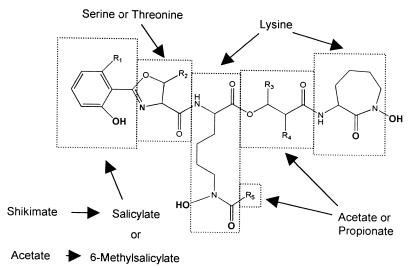
General structure of MBs. Iron-chelating groups are in boldface. Letters and R groups correspond to the structures listed in Fig. 3.
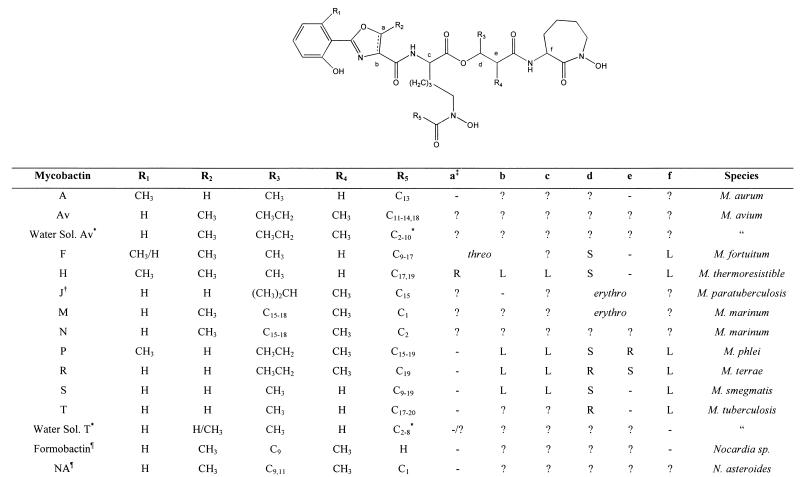
General structures of known MBs. Symbols: ∗, the terminal carbon in this chain of this water-soluble MB is either a carboxylic acid or its methyl ester; †, the structure as originally reported (49); §, MB that contains an oxazole moiety instead of the usual oxazoline and is found in Nocardia sp; ‡, letters refer to the stereocenters shown in the structure above, − indicates that the center is not chiral in this compound, and ? indicates that the stereochemistry is unknown.
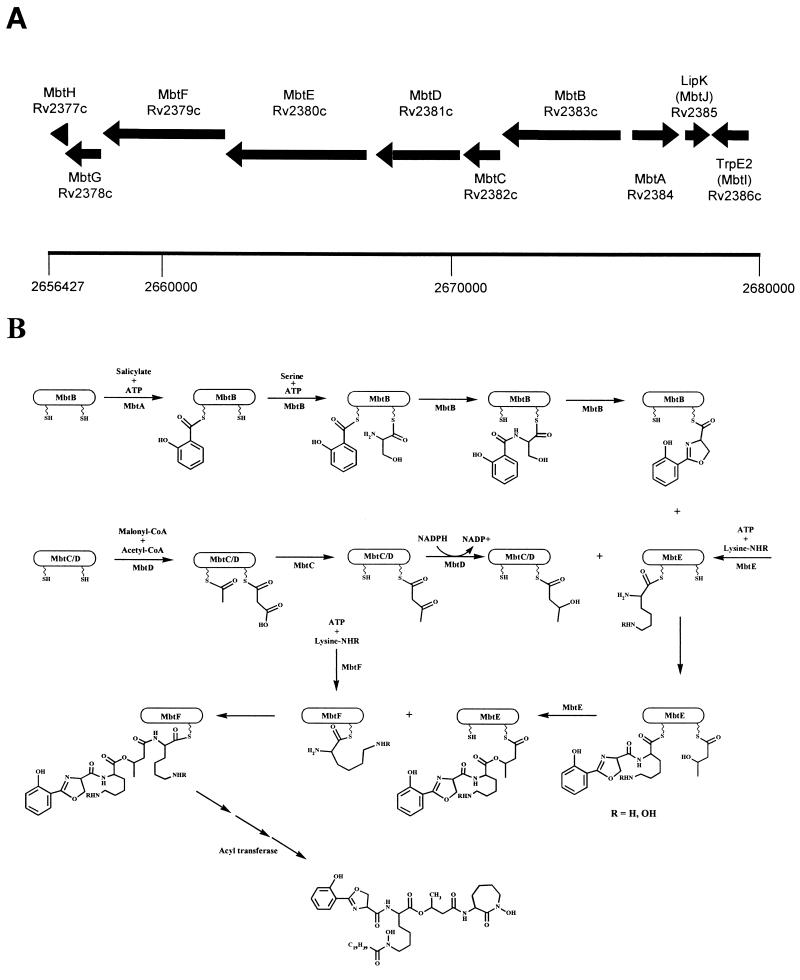
Proposed biosynthetic scheme for MB T. (A) Region of the genome of M. tuberculosis H37Rv containing the putative MB biosynthetic enzymes. Numbering refers to chromosomal positions reported by Cole et al. (8). (B) Proposed biosynthetic cascade catalyzed by the Mbt locus. See the text for details of individual enzymatic predictions. There are significant areas of uncertainty in this model, as suggested by the following questions. (i) Does the ketoacyl synthase domain of MbtC act upon the MbtD-bound acyl carrier protein domains as shown? (ii) Does the second peptidyl/acyl carrier protein domain of MbtE function to attach the β-hydroxybutyrate as shown, or does it transfer the acyl group to the first lysine (the R5) position in Fig. 2? (iii) Do MbtE and MbtF act in the order in which they appear in the genomic locus, or are they reversed? (iv) Is MbtG involved in N-hydroxylation as shown? (v) What is the nature of the acyltransferase, and how is the specificity determined? When does this act and upon what substrate? Since this determines the ratio of water-soluble to cell-associated siderophore, this is a critical question. (vi) What is the role played by MbtH and MbtJ?
References
-
- Allen M, Birch A J, Jones A R. Studies in relation to biosynthesis. XLIII. Incorporation of l-lysine into mycobactin-P. Aust J Chem. 1970;23:427–429.
-
- Andrews S C. Iron storage in bacteria. Adv Microb Physiol. 1998;40:281–351. - PubMed
-
- Baynes R D, Flax H, Bothwell T H, Bezwoda W R, MacPhail A P, Atkinson P, Lewis D. Haematological and iron-related measurements in active pulmonary tuberculosis. Scand J Haematol. 1986;36:280–287. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
