

Targeted disruption of Fgf8 causes failure of cell migration in the gastrulating mouse embryo
- PMID: 10421635
- PMCID: PMC316887
- DOI: 10.1101/gad.13.14.1834
Targeted disruption of Fgf8 causes failure of cell migration in the gastrulating mouse embryo
Abstract
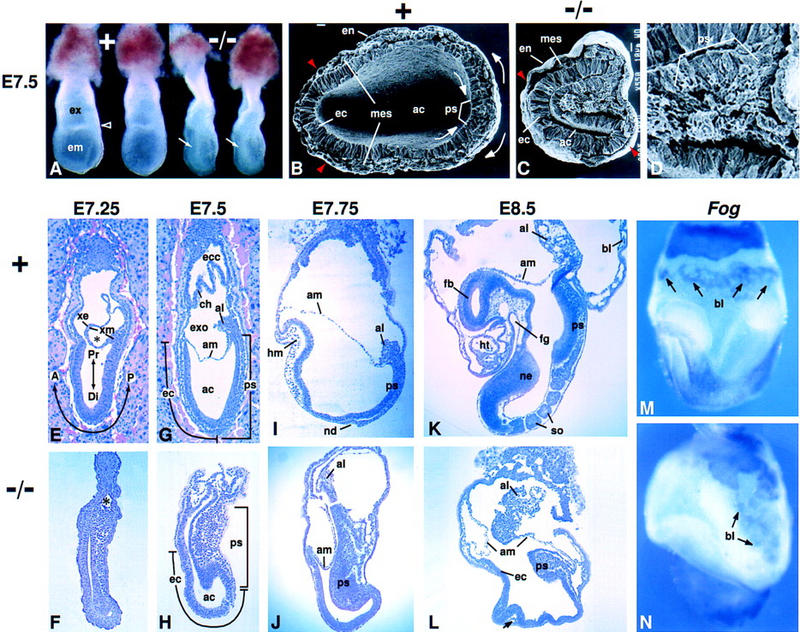
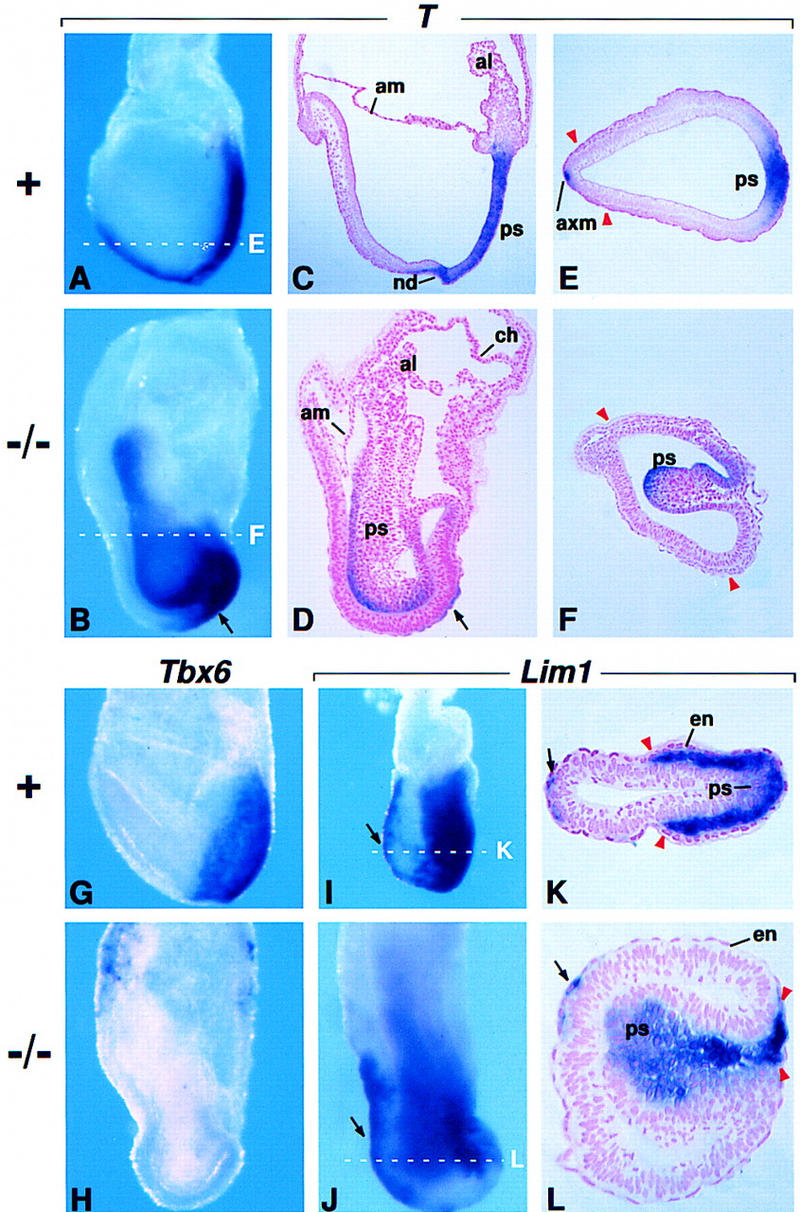
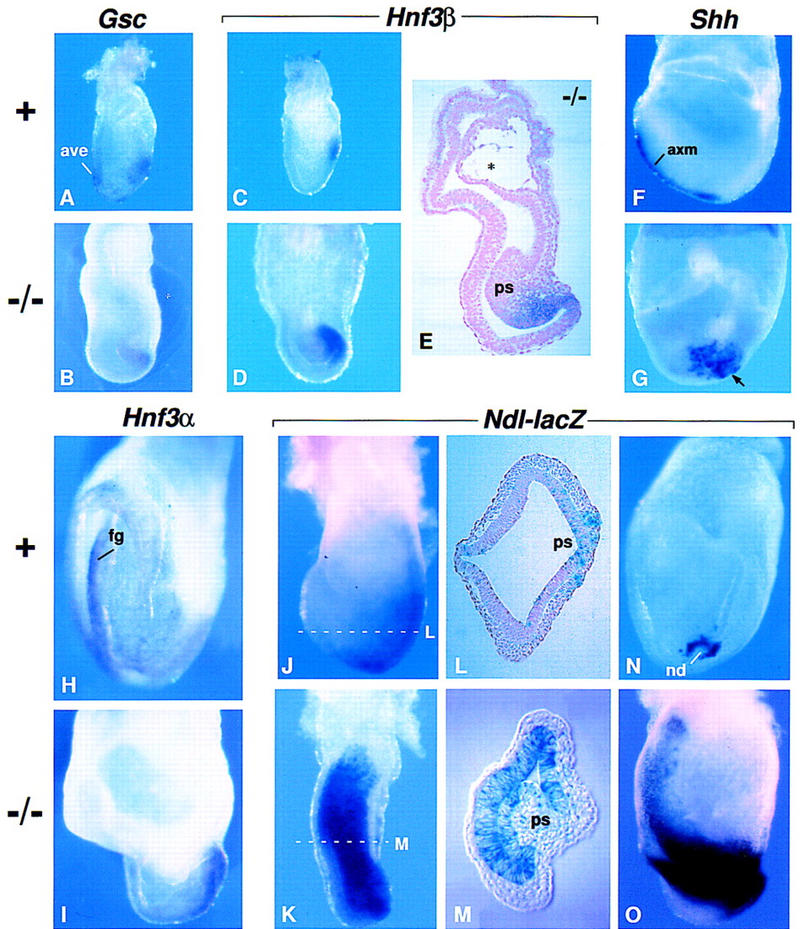
Fgf8 and Fgf4 encode FGF family members that are coexpressed in the primitive streak of the gastrulating mouse embryo. We have analyzed the phenotype of Fgf8(-/-) embryos and discovered that they fail to express Fgf4 in the streak. In the absence of both FGF8 and FGF4, epiblast cells move into the streak and undergo an epithelial-to-mesenchymal transition, but most cells then fail to move away from the streak. As a consequence, no embryonic mesoderm- or endoderm-derived tissues develop, although extraembryonic tissues form. Patterning of the prospective neuroectoderm is greatly perturbed in the mutant embryos. Anterior neuroectoderm markers are widely expressed, at least in part because the anterior visceral endoderm, which provides signals that regulate their expression, is not displaced proximally in the absence of definitive endoderm. Posterior neuroectoderm markers are not expressed, presumably because there is neither mesendoderm underlying the prospective neuroectoderm nor a morphologically normal node to provide the inductive signals necessary for their expression. This study identifies Fgf8 as a gene essential for gastrulation and shows that signaling via FGF8 and/or FGF4 is required for cell migration away from the primitive streak.
Figures


Similar articles
-
Cell movement patterns during gastrulation in the chick are controlled by positive and negative chemotaxis mediated by FGF4 and FGF8.Dev Cell. 2002 Sep;3(3):425-37. doi: 10.1016/s1534-5807(02)00256-3. Dev Cell. 2002. PMID: 12361604
-
The Wnt co-receptors Lrp5 and Lrp6 are essential for gastrulation in mice.Development. 2004 Jun;131(12):2803-15. doi: 10.1242/dev.01137. Epub 2004 May 13. Development. 2004. PMID: 15142971
-
Hypoblast controls mesoderm generation and axial patterning in the gastrulating rabbit embryo.Dev Genes Evol. 2004 Dec;214(12):591-605. doi: 10.1007/s00427-004-0436-y. Epub 2004 Oct 6. Dev Genes Evol. 2004. PMID: 15480760
-
Establishment and organization of germ layers in the gastrulating mouse embryo.Ciba Found Symp. 1992;165:27-41; discussion 42-9. doi: 10.1002/9780470514221.ch3. Ciba Found Symp. 1992. PMID: 1516473 Review.
-
The mouse fetal-placental arterial connection: A paradigm involving the primitive streak and visceral endoderm with implications for human development.Wiley Interdiscip Rev Dev Biol. 2020 Mar;9(2):e362. doi: 10.1002/wdev.362. Epub 2019 Oct 17. Wiley Interdiscip Rev Dev Biol. 2020. PMID: 31622045 Review.
Cited by
-
Mechanical Coupling Coordinates the Co-elongation of Axial and Paraxial Tissues in Avian Embryos.Dev Cell. 2020 Nov 9;55(3):354-366.e5. doi: 10.1016/j.devcel.2020.08.007. Epub 2020 Sep 11. Dev Cell. 2020. PMID: 32918876 Free PMC article.
-
Fgf regulates dedifferentiation during skeletal muscle regeneration in adult zebrafish.Cell Signal. 2016 Sep;28(9):1196-1204. doi: 10.1016/j.cellsig.2016.06.001. Epub 2016 Jun 3. Cell Signal. 2016. PMID: 27267062 Free PMC article.
-
Sonic hedgehog protein regulates fibroblast growth factor 8 expression in metanephric explant culture from BALB/c mice: Possible mechanisms associated with renal morphogenesis.Mol Med Rep. 2016 Oct;14(4):2929-36. doi: 10.3892/mmr.2016.5614. Epub 2016 Aug 9. Mol Med Rep. 2016. PMID: 27510750 Free PMC article.
-
Evolution of vertebrate forebrain development: how many different mechanisms?J Anat. 2001 Jul-Aug;199(Pt 1-2):35-52. doi: 10.1046/j.1469-7580.2001.19910035.x. J Anat. 2001. PMID: 11523828 Free PMC article. Review.
-
Tbx1 and Foxi3 genetically interact in the pharyngeal pouch endoderm in a mouse model for 22q11.2 deletion syndrome.PLoS Genet. 2019 Aug 14;15(8):e1008301. doi: 10.1371/journal.pgen.1008301. eCollection 2019 Aug. PLoS Genet. 2019. PMID: 31412026 Free PMC article.
References
-
- Amaya E, Musci TJ, Kirschner MW. Expression of a dominant negative mutant of the FGF receptor disrupts mesoderm formation in Xenopus embryos. Cell. 1991;66:257–270. - PubMed
-
- Ang S-L, Wierda A, Wong D, Stevens KA, Cascio S, Rossant J, Zaret KS. The formation and maintenance of the definitive endoderm lineage in the mouse: involvement of HNF3/forkhead proteins. Development. 1993;119:1301–1315. - PubMed
-
- Ang S-L, Conlon RA, Jin O, Rossant J. Positive and negative signals from mesoderm regulate the expression of mouse Otx2 in ectoderm explants. Development. 1994;120:2979–2989. - PubMed
-
- Baldwin HS, Shen HM, Yan HC, DeLisser HM, Chung A, Mickanin C, Trask T, Kirschbaum NE, Newman PJ, Albelda SM, Buck CA. Platelet endothelial cell adhesion molecule-1 (PECAM-1/CD31): Alternatively spliced, functionally distinct isoforms expressed during mammalian cardiovascular development. Development. 1994;120:2539–2553. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials