

Collisions between yeast chromosomal loci in vivo are governed by three layers of organization
- PMID: 10421638
- PMCID: PMC316882
- DOI: 10.1101/gad.13.14.1871
Collisions between yeast chromosomal loci in vivo are governed by three layers of organization
Abstract
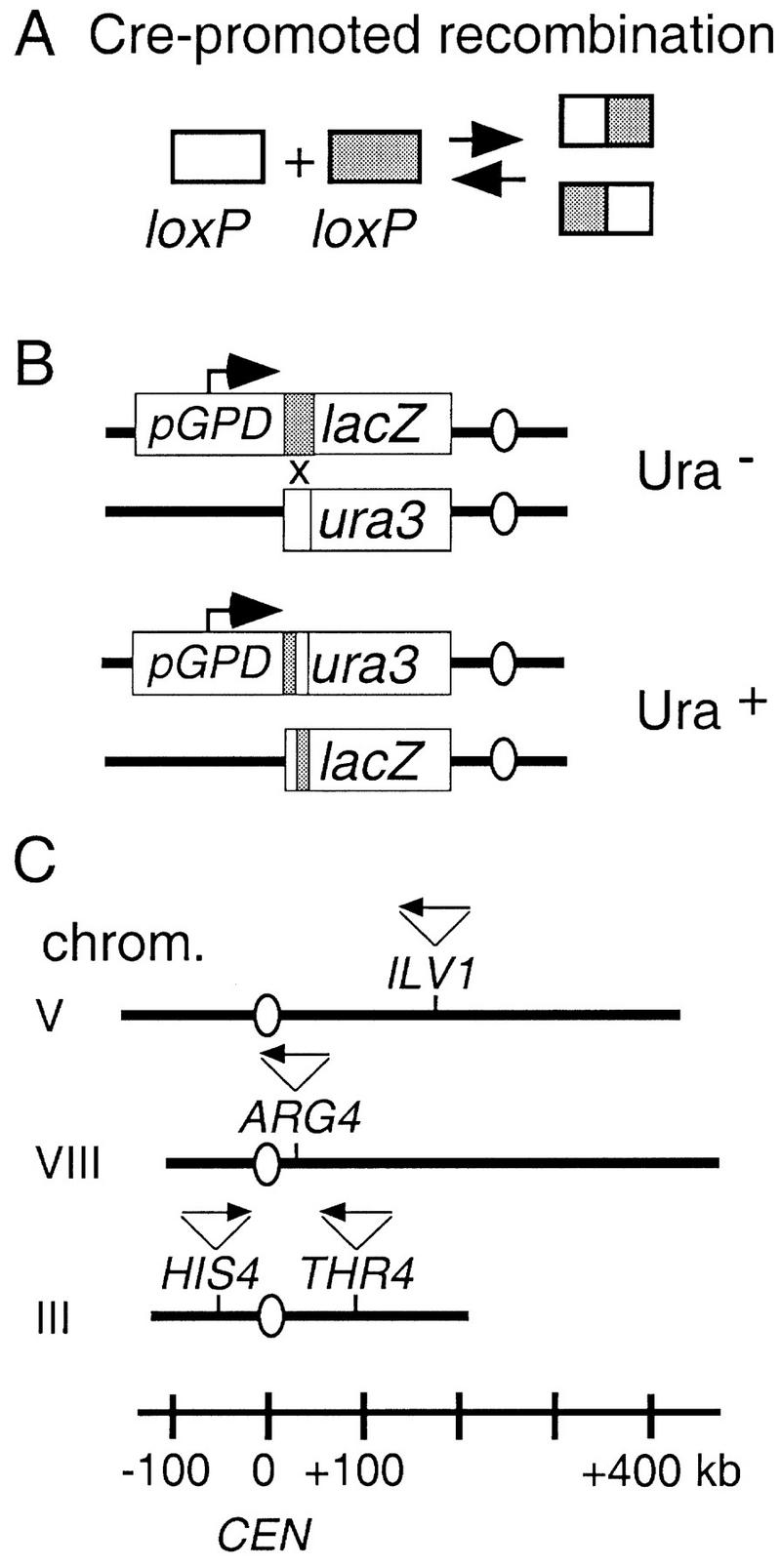
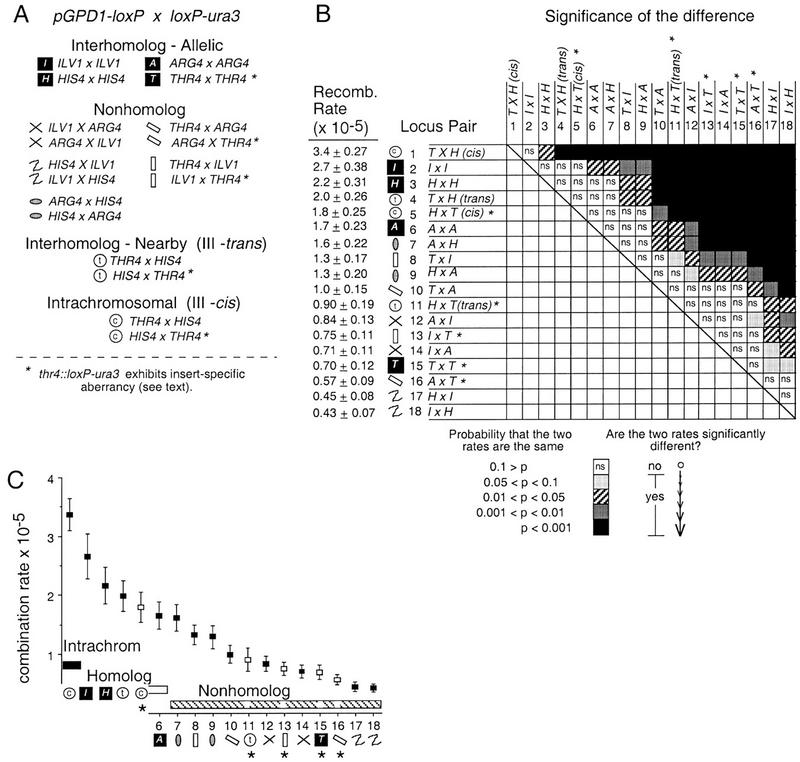
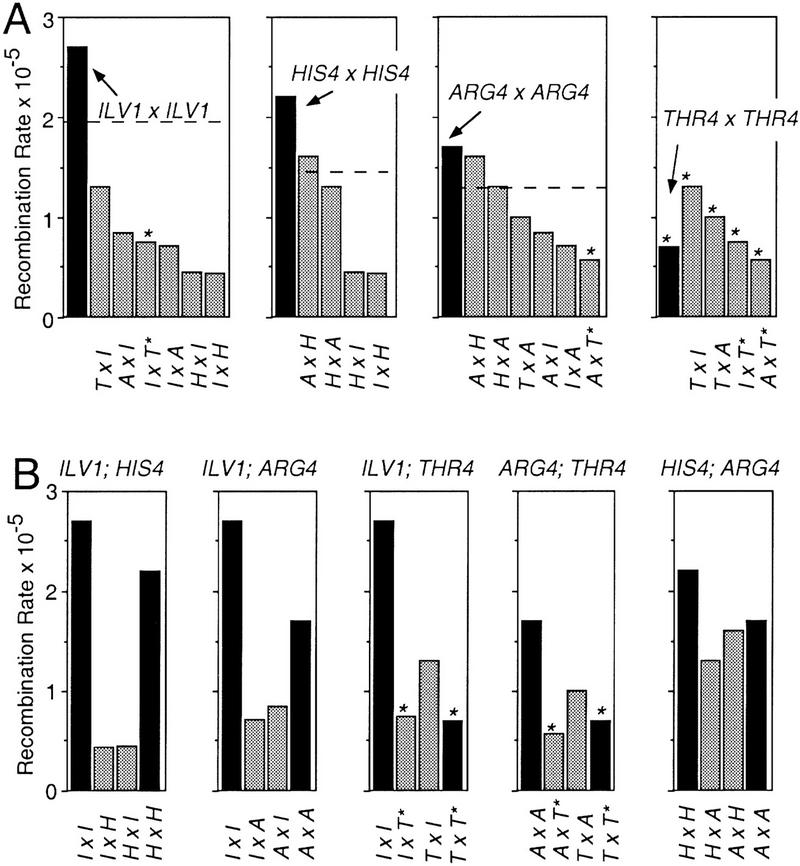
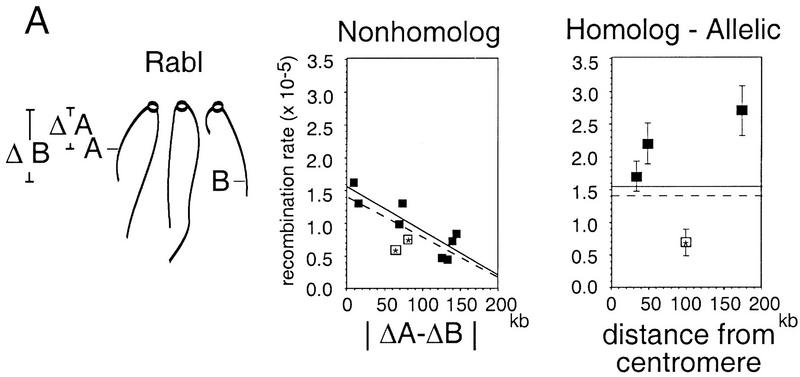
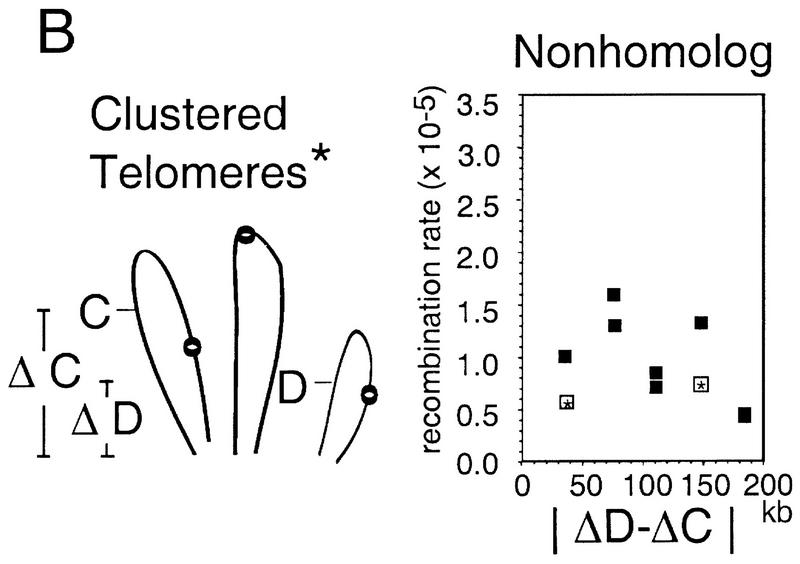
The relative probabilities that different pairs of chromosomal loci will collide with one another in vegetatively growing diploid yeast cells have been assessed using a genetic assay for Cre/loxP site-specific recombination. Recombination rates have been determined for 18 different pairs of loxP sites representing diverse pairs of positions within the genome. Overall, relative collision probabilities vary over an eightfold range. Within this range, a hierarchy comprising three levels of organization can be discerned. First, collisions between loci on nonhomologous chromosomes are governed by nonspecific centromere clustering. Second, a sequence is closer to allelic or nearby sequences on its homolog than to sequences on nonhomologous chromosomes, an effect most simply attributed to homolog pairing. Third, a sequence can be closer to other sequences nearby on the same chromosome than to sequences on other chromosomes. These findings provide a framework for assessing the role of chromosome disposition in cellular processes such as DNA repair and gene expression. Also the possibility is raised that genome-wide coalignment of homologs is not the fundamental raison d'etre of the somatic pairing process. We suggest instead that pairing may exist to promote juxtaposition of homologous regions within irregular genome complements.
Figures
References
-
- Ambremski K, Hoess R, Sternberg N. Studies on the properties of P1 site-specific recombination: Evidence for topologically unlinked products following recombination. Cell. 1983;32:1301–1311. - PubMed
-
- Bender JC. Cytosine methylation of repeated sequences in eukaryotes: The role of DNA pairing. Trends Biochem Sci. 1998;23:252–256. - PubMed
-
- Berben G, Dumont J, Gilliquet V, Bolle PA, Hilger F. The YDp plasmids: A uniform set of vectors bearing versatile gene disruption cassettes for Saccharomyces cerevisiae. Yeast. 1991;7:475–477. - PubMed
-
- Bliska JB, Cozzarelli NR. Use of site-specific recombination as a probe of DNA structure and metabolism in vivo. J Mol Biol. 1987;194:205–218. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases