

Establishment of the dorsal-ventral axis in Xenopus embryos coincides with the dorsal enrichment of dishevelled that is dependent on cortical rotation
- PMID: 10427095
- PMCID: PMC2156185
- DOI: 10.1083/jcb.146.2.427
Establishment of the dorsal-ventral axis in Xenopus embryos coincides with the dorsal enrichment of dishevelled that is dependent on cortical rotation
Abstract
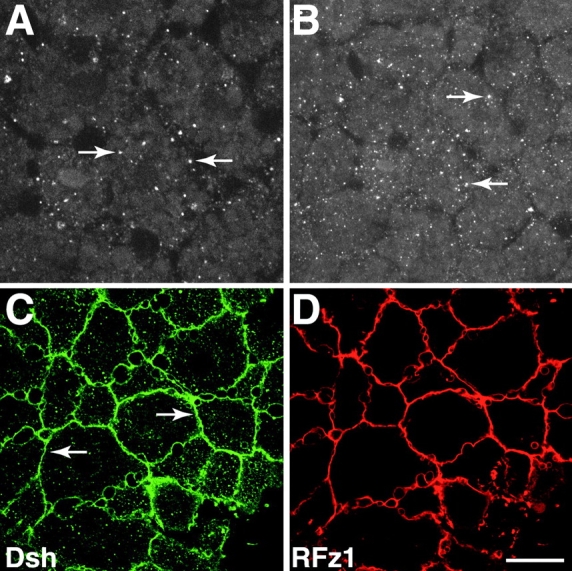
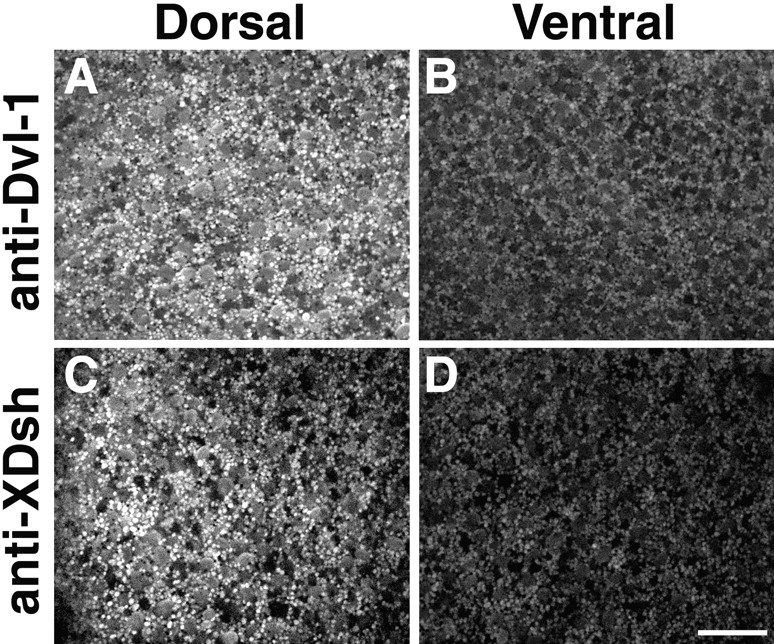
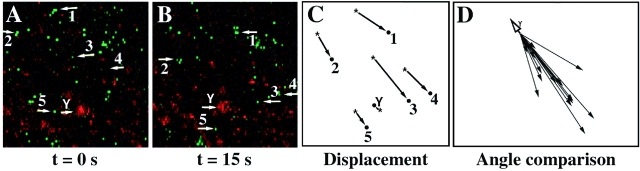
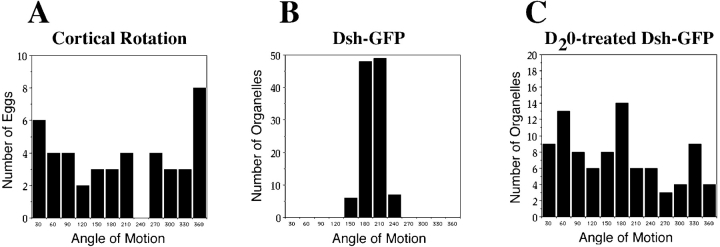
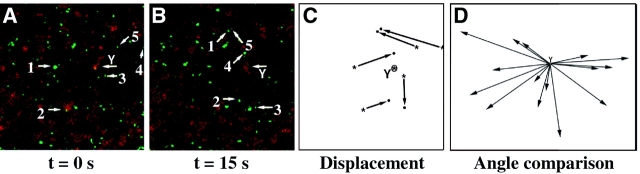
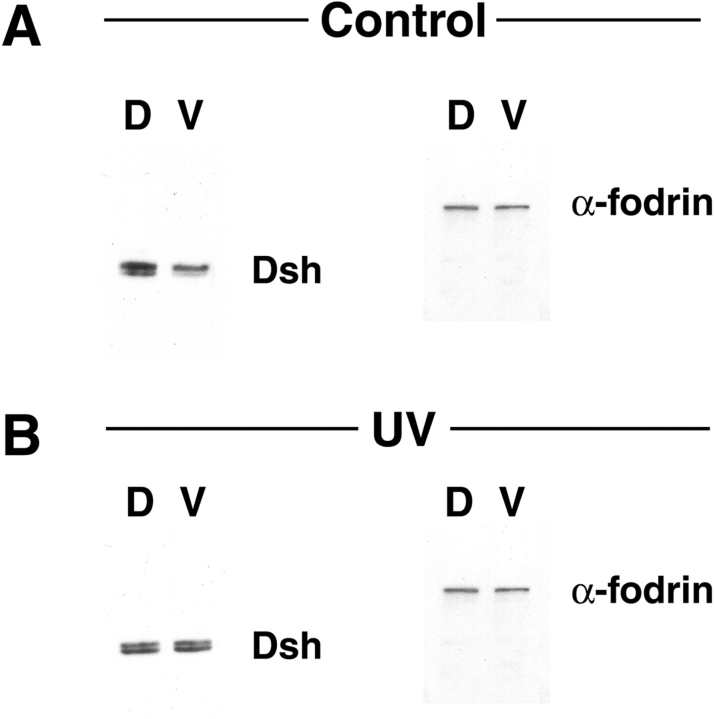
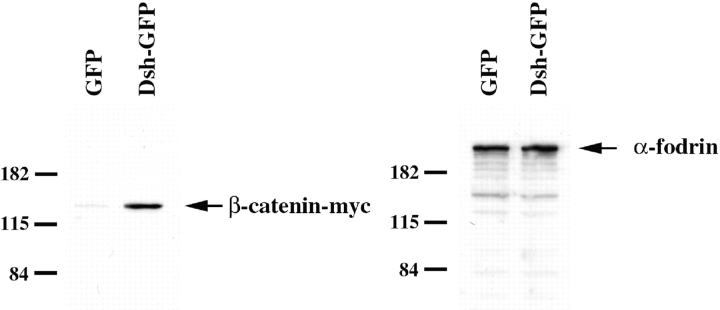
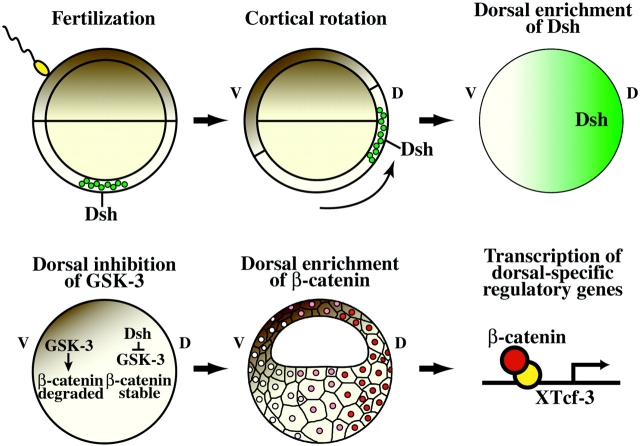
Examination of the subcellular localization of Dishevelled (Dsh) in fertilized Xenopus eggs revealed that Dsh is associated with vesicle-like organelles that are enriched on the prospective dorsal side of the embryo after cortical rotation. Dorsal enrichment of Dsh is blocked by UV irradiation of the vegetal pole, a treatment that inhibits development of dorsal cell fates, linking accumulation of Dsh and specification of dorsal cell fates. Investigation of the dynamics of Dsh localization using Dsh tagged with green fluorescent protein (Dsh-GFP) demonstrated that Dsh-GFP associates with small vesicle-like organelles that are directionally transported along the parallel array of microtubules towards the prospective dorsal side of the embryo during cortical rotation. Perturbing the assembly of the microtubule array with D(2)O, a treatment that promotes the random assembly of the array and the dorsalization of embryos, randomizes translocation of Dsh-GFP. Conversely, UV irradiation of the vegetal pole abolishes movement of Dsh-GFP. Finally, we demonstrate that overexpression of Dsh can stabilize beta-catenin in Xenopus. These data suggest that the directional translocation of Dsh along microtubules during cortical rotation and its subsequent enrichment on the prospective dorsal side of the embryo play a role in locally activating a maternal Wnt pathway responsible for establishing dorsal cell fates in Xenopus.
Figures
Similar articles
-
Microtubule-mediated transport of organelles and localization of beta-catenin to the future dorsal side of Xenopus eggs.Proc Natl Acad Sci U S A. 1997 Feb 18;94(4):1224-9. doi: 10.1073/pnas.94.4.1224. Proc Natl Acad Sci U S A. 1997. PMID: 9037034 Free PMC article.
-
Rap2 is required for Wnt/beta-catenin signaling pathway in Xenopus early development.EMBO J. 2005 Mar 9;24(5):985-96. doi: 10.1038/sj.emboj.7600571. Epub 2005 Feb 10. EMBO J. 2005. PMID: 15706349 Free PMC article.
-
Move it or lose it: axis specification in Xenopus.Development. 2004 Aug;131(15):3491-9. doi: 10.1242/dev.01284. Development. 2004. PMID: 15262887 Review.
-
Differential stability of beta-catenin along the animal-vegetal axis of the sea urchin embryo mediated by dishevelled.Development. 2004 Jun;131(12):2947-56. doi: 10.1242/dev.01152. Epub 2004 May 19. Development. 2004. PMID: 15151983
-
New steps in the Wnt/beta-catenin signal transduction pathway.Recent Prog Horm Res. 2000;55:225-36. Recent Prog Horm Res. 2000. PMID: 11036939 Review.
Cited by
-
Autoinhibition of Dishevelled protein regulated by its extreme C terminus plays a distinct role in Wnt/β-catenin and Wnt/planar cell polarity (PCP) signaling pathways.J Biol Chem. 2017 Apr 7;292(14):5898-5908. doi: 10.1074/jbc.M116.772509. Epub 2017 Feb 21. J Biol Chem. 2017. PMID: 28223363 Free PMC article.
-
Vegetally localized Xenopus trim36 regulates cortical rotation and dorsal axis formation.Development. 2009 Sep;136(18):3057-65. doi: 10.1242/dev.036855. Epub 2009 Aug 12. Development. 2009. PMID: 19675128 Free PMC article.
-
Molecular specification of germ layers in vertebrate embryos.Cell Mol Life Sci. 2016 Mar;73(5):923-47. doi: 10.1007/s00018-015-2092-y. Epub 2015 Dec 14. Cell Mol Life Sci. 2016. PMID: 26667903 Free PMC article. Review.
-
Maternal Dead-End1 is required for vegetal cortical microtubule assembly during Xenopus axis specification.Development. 2013 Jun;140(11):2334-44. doi: 10.1242/dev.094748. Epub 2013 Apr 24. Development. 2013. PMID: 23615278 Free PMC article.
-
Wnt signalling antagonizes stress granule assembly through a Dishevelled-dependent mechanism.Biol Open. 2012 Feb 15;1(2):109-19. doi: 10.1242/bio.2011023. Epub 2011 Nov 18. Biol Open. 2012. PMID: 23213403 Free PMC article.
References
-
- Axelrod J.D., Matsuno K., Artavanis-Tsakonas S., Perrimon N. Interaction between Wingless and Notch signaling pathways mediated by Dishevelled. Science. 1996;271:1826–1832. - PubMed
-
- Brannon M., Brown J.D., Bates R., Kimelman D., Moon R.T. XCtBP is a XTcf-3 co-repressor with roles throughout Xenopus development. Development. 1999;126:3159–3170. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
