

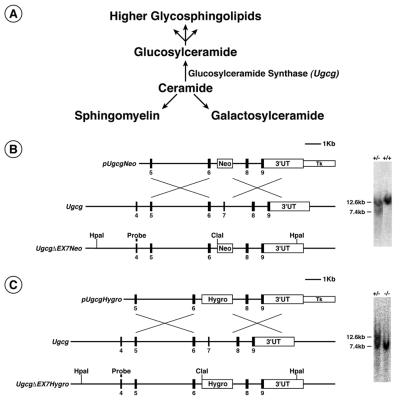
A vital role for glycosphingolipid synthesis during development and differentiation
- PMID: 10430909
- PMCID: PMC17746
- DOI: 10.1073/pnas.96.16.9142
A vital role for glycosphingolipid synthesis during development and differentiation
Abstract
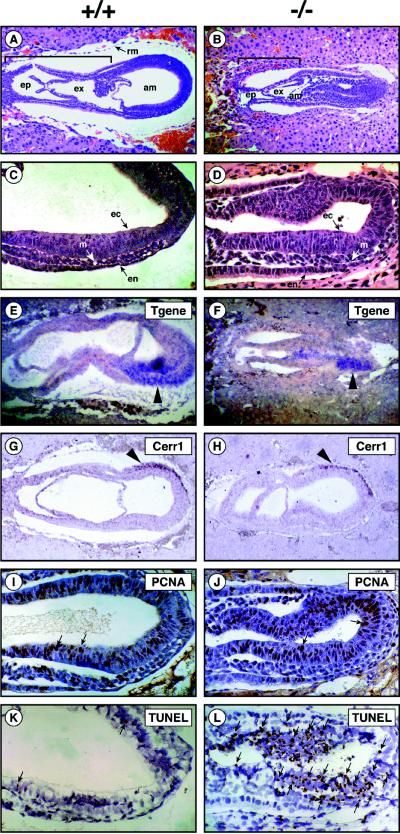
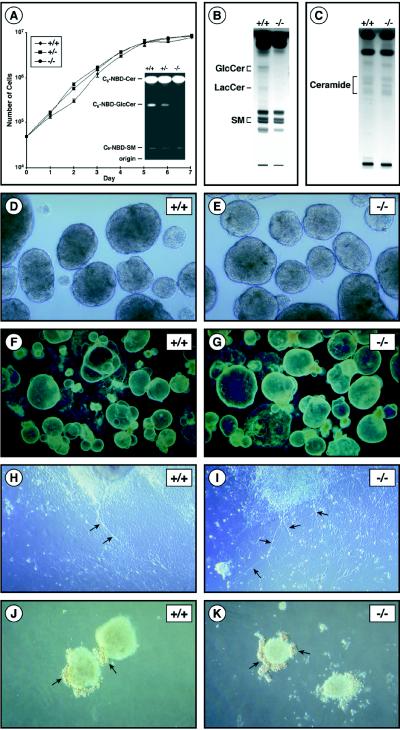
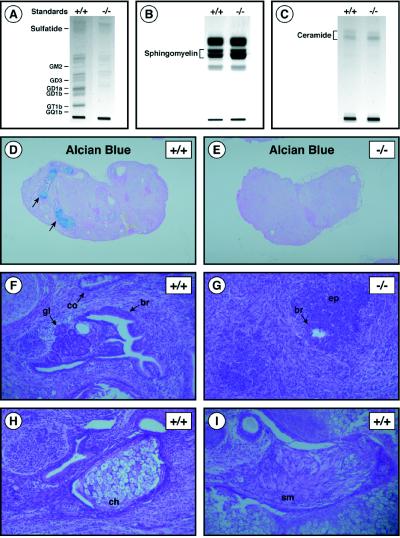
Glycosphingolipids (GSLs) are believed to be integral for the dynamics of many cell membrane events, including cellular interactions, signaling, and trafficking. We have investigated their roles in development and differentiation by eliminating the major synthesis pathway of GSLs through targeted disruption of the Ugcg gene encoding glucosylceramide synthase. In the absence of GSL synthesis, embryogenesis proceeded well into gastrulation with differentiation into primitive germ layers and patterning of the embryo but was abruptly halted by a major apoptotic process. In vivo, embryonic stem cells deficient in GSL synthesis were again able to differentiate into endodermal, mesodermal, and ectodermal derivatives but were strikingly deficient in their ability to form well differentiated tissues. In vitro, however, hematopoietic and neuronal differentiation could be induced. The results demonstrate that the synthesis of GSL structures is essential for embryonic development and for the differentiation of some tissues and support the concept that GSLs are involved in crucial cell interactions mediating these processes.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
