

The Epstein-Barr virus latency BamHI-Q promoter is positively regulated by STATs and Zta interference with JAK/STAT activation leads to loss of BamHI-Q promoter activity
- PMID: 10430944
- PMCID: PMC17784
- DOI: 10.1073/pnas.96.16.9339
The Epstein-Barr virus latency BamHI-Q promoter is positively regulated by STATs and Zta interference with JAK/STAT activation leads to loss of BamHI-Q promoter activity
Abstract
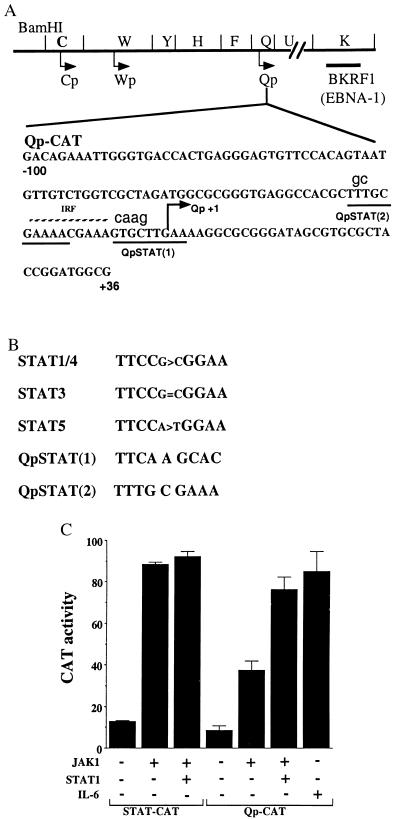
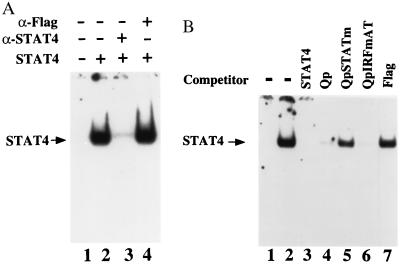
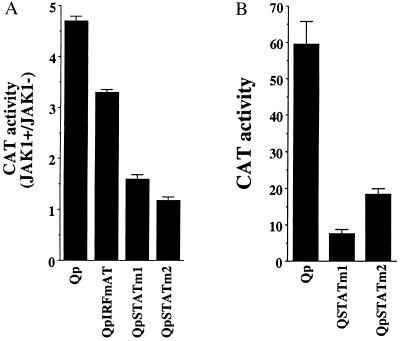
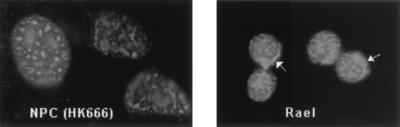
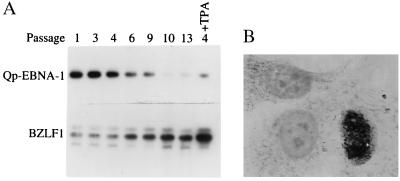
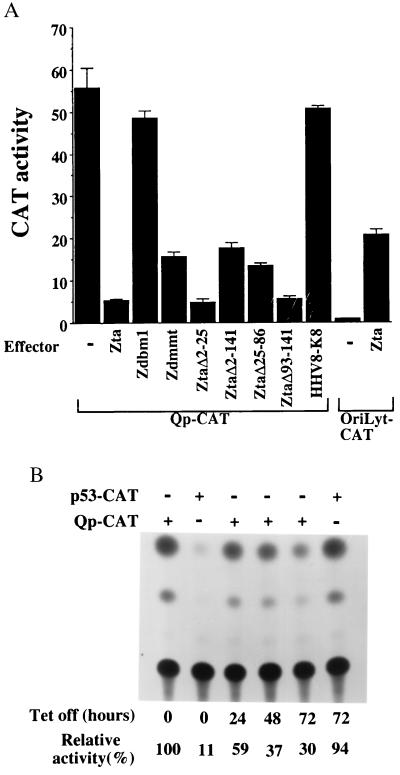
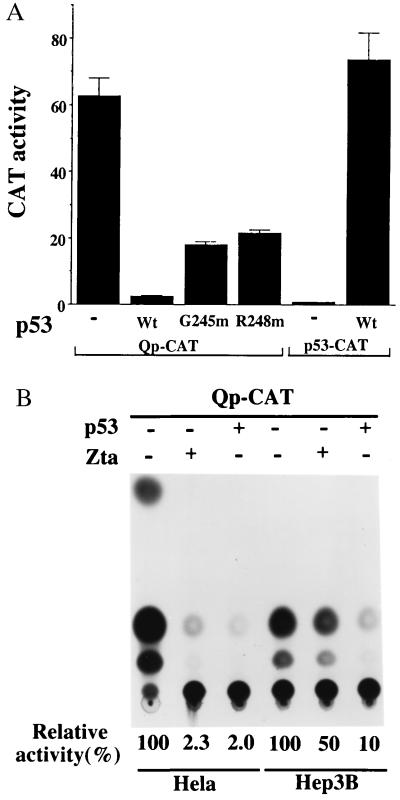
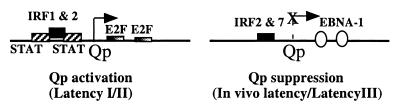
In Epstein-Barr virus (EBV)-associated tumors in nonimmunocompromised patients, EBV gene expression is highly restricted. EBV-encoded nuclear antigen (EBNA)-1 is expressed, whereas the immunogenic and proliferative EBNAs are not. This pattern of EBNA expression is generated by usage of the BamHI-Q promoter (Qp). We have determined that the JAK/STAT pathway positively regulates Qp activity. In transient-transfection assays, a Qp-CAT reporter was activated by cotransfected JAK-1 and by treatment of cells with the cytokine IL-6. The ability of Qp to bind signal transducer and activator of transcription (STAT) proteins was directly demonstrated by electrophoretic mobility-shift assay, and mutation of potential STAT-binding sites reduced Qp responsiveness to Janus kinase (JAK)-1. Consistent with a role for STATs in Qp function, Qp using Burkitt's lymphoma Rael cells and cultured nasopharyngeal carcinoma (NPC) cells contained nuclear STAT protein. We investigated whether the inability to maintain EBV-positive NPC cell lines in culture was related to Qp activity. Passaging of the NPC cell line HK666 led to activation of expression of BZLF1, which encodes Zta and loss of Qp function. Transient expression assays linked Zta expression to the down-regulation of Qp. Cotransfection of Zta reduced Qp activity in reporter assays. This negative regulation required Zta DNA-binding activity. We provide evidence that Zta up-regulation of p53 leads to p53-mediated interference with JAK/STAT activation of Qp. The data imply that JAK/STAT signaling has a role in EBV-associated malignancies.
Figures

References
-
- Rickinson A B, Kieff E. In: Field’s Virology. Field B N, Knipe D M, Howley P M, editors. Vol. 2. New York: Raven; 1996. pp. 2397–2446.
-
- Kieff E. In: Field’s Virology. Field B N, Knipe D M, Howley P M, editors. Vol. 2. New York: Raven; 1996. pp. 2343–2396.
-
- Rickinson A B, Moss D J. Annu Rev Immunol. 1997;15:405–431. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
