

A novel H(+) conductance in eosinophils: unique characteristics and absence in chronic granulomatous disease
- PMID: 10432282
- PMCID: PMC2195580
- DOI: 10.1084/jem.190.2.183
A novel H(+) conductance in eosinophils: unique characteristics and absence in chronic granulomatous disease
Abstract
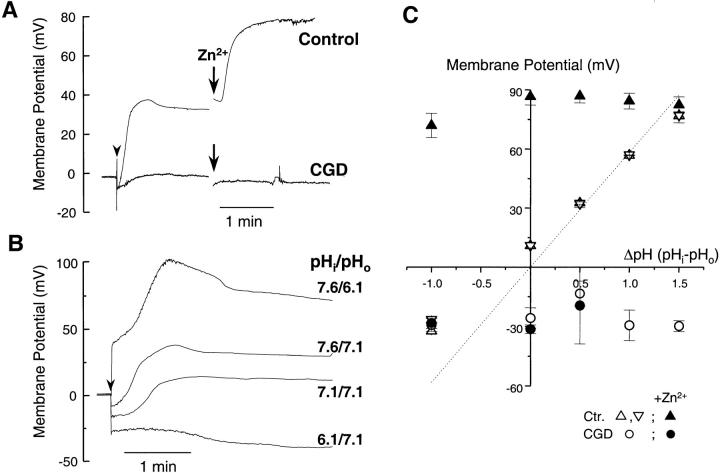
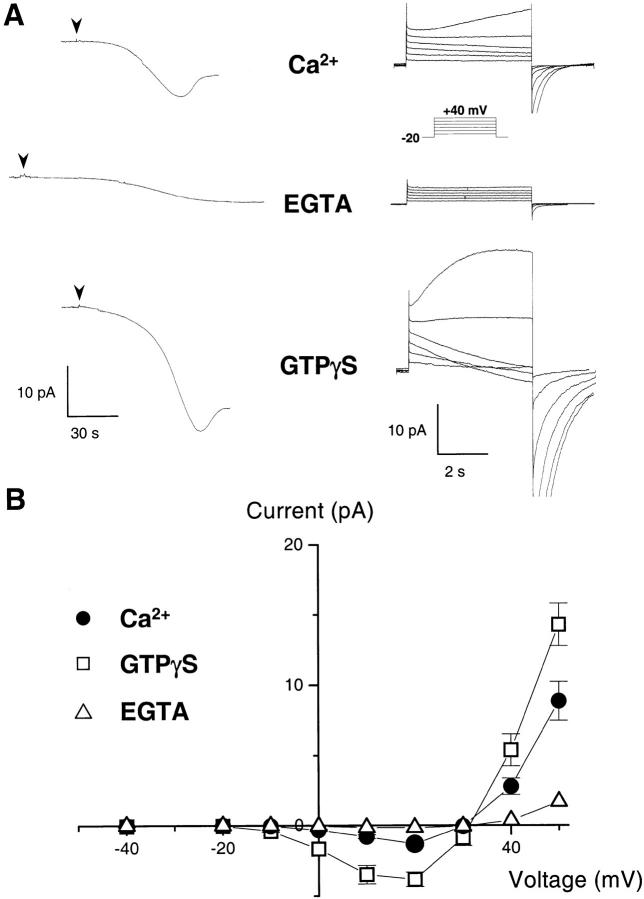
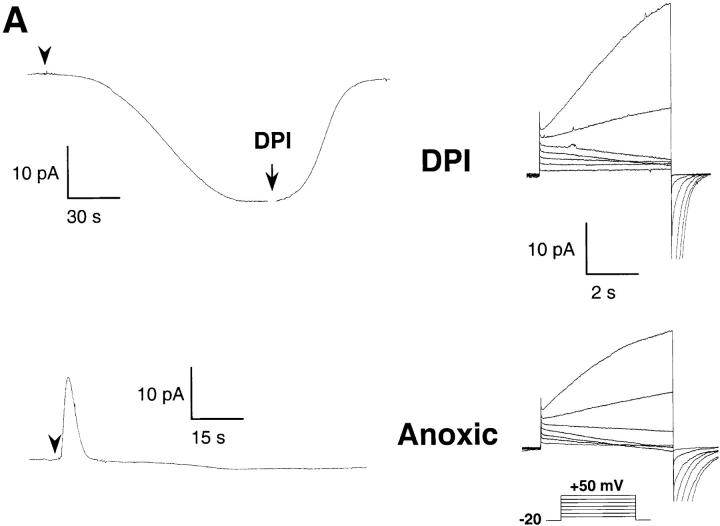
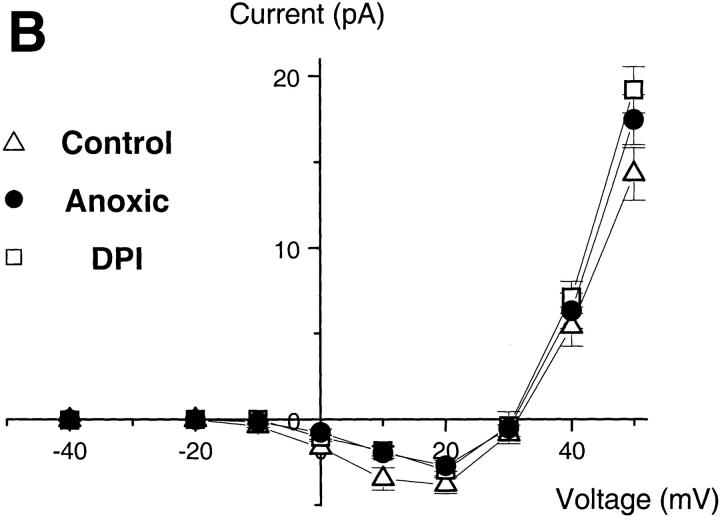
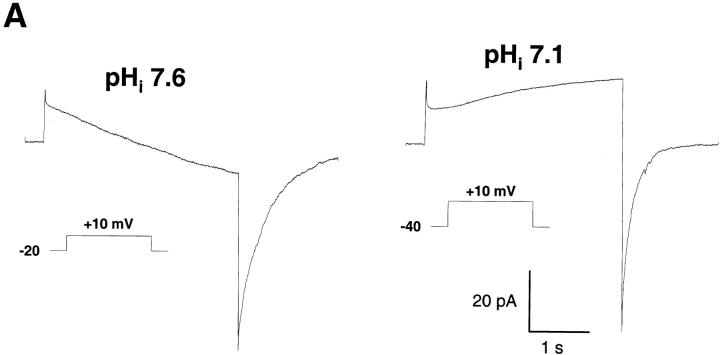
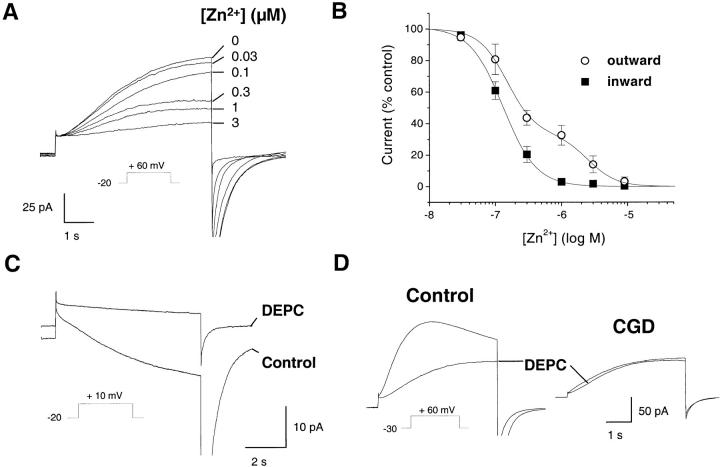
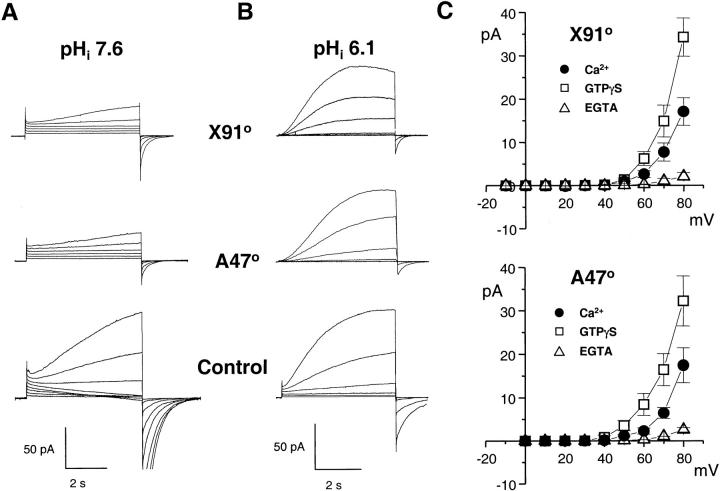
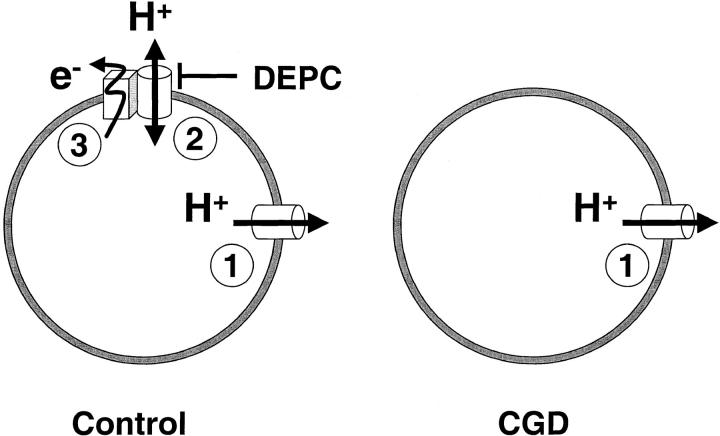
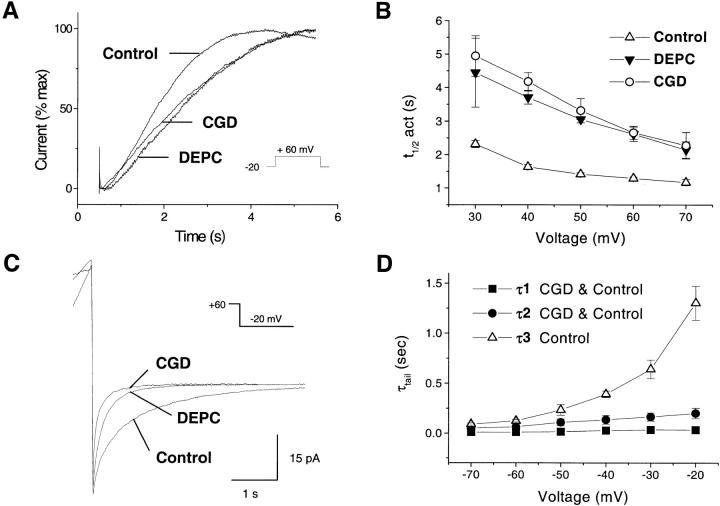
Efficient mechanisms of H(+) ion extrusion are crucial for normal NADPH oxidase function. However, whether the NADPH oxidase-in analogy with mitochondrial cytochromes-has an inherent H(+) channel activity remains uncertain: electrophysiological studies did not find altered H(+) currents in cells from patients with chronic granulomatous disease (CGD), challenging earlier reports in intact cells. In this study, we describe the presence of two different types of H(+) currents in human eosinophils. The "classical" H(+) current had properties similar to previously described H(+) conductances and was present in CGD cells. In contrast, the "novel" type of H(+) current had not been described previously and displayed unique properties: (a) it was absent in cells from gp91- or p47-deficient CGD patients; (b) it was only observed under experimental conditions that allowed NADPH oxidase activation; (c) because of its low threshold of voltage activation, it allowed proton influx and cytosolic acidification; (d) it activated faster and deactivated with slower and distinct kinetics than the classical H(+) currents; and (e) it was approximately 20-fold more sensitive to Zn(2+) and was blocked by the histidine-reactive agent, diethylpyrocarbonate (DEPC). In summary, our results demonstrate that the NADPH oxidase or a closely associated protein provides a novel type of H(+) conductance during phagocyte activation. The unique properties of this conductance suggest that its physiological function is not restricted to H(+) extrusion and repolarization, but might include depolarization, pH-dependent signal termination, and determination of the phagosomal pH set point.
Figures


References
-
- Babior B.M. Oxygen-dependent microbial killing by phagocytes (first of two parts) N. Engl. J. Med. 1978;298:659–668. - PubMed
-
- Klebanoff S.J. Oxygen metabolism and the toxic properties of phagocytes. Ann. Intern. Med. 1980;93:480–489. - PubMed
-
- Segal A.W. The antimicrobial role of the neutrophil leukocyte. J. Infect. 1981;3:3–17. - PubMed
-
- Clark R.A. The human neutrophil respiratory burst oxidase. J. Infect. Dis. 1990;161:1140–1147. - PubMed
-
- Segal A.W., Abo A. The biochemical basis of the NADPH oxidase of phagocytes. Trends Biochem. Sci. 1993;18:43–47. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
