

Nuclear export factor CRM1 interacts with nonstructural proteins NS2 from parvovirus minute virus of mice
- PMID: 10438867
- PMCID: PMC104304
- DOI: 10.1128/JVI.73.9.7769-7779.1999
Nuclear export factor CRM1 interacts with nonstructural proteins NS2 from parvovirus minute virus of mice
Abstract
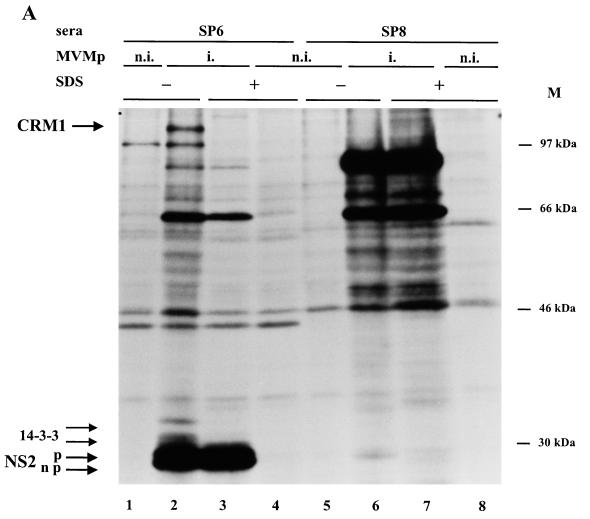
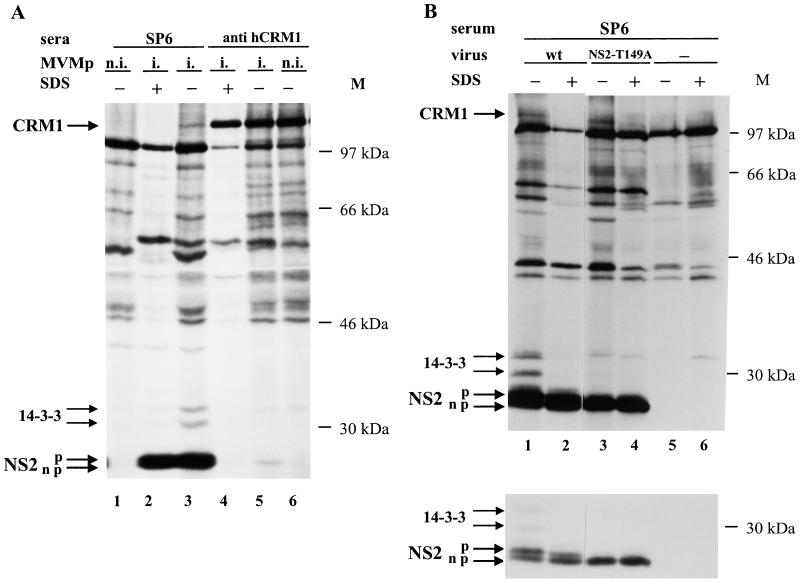
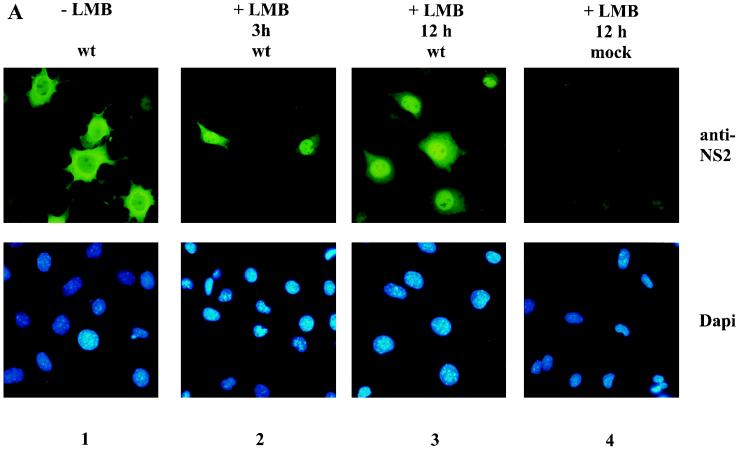
The nonstructural NS2 proteins of autonomous parvoviruses are known to act in a host cell-dependent manner and to play a role in viral DNA replication, efficient translation of viral mRNA, and/or encapsidation. Their exact function during the parvovirus life cycle remains, however, still obscure. We report here the characterization of the interaction with the NS2 proteins from the parvovirus minute virus of mice (MVM) and rat as well as mouse homologues of the human CRM1 protein, a member of the importin-beta family recently identified as an essential nuclear export factor. Using the two-hybrid system, we could detect the interaction between the carboxy-terminal region of rat CRM1 and each of the three isoforms of NS2 (P [or major], Y [or minor], and L [or rare]). NS2 proteins were further shown to interact with the full-length CRM1 by coimmunoprecipitation experiments using extracts from both mouse and rat cell lines. Our data show that CRM1 preferentially binds to the nonphosphorylated isoforms of NS2. Moreover, we observed that the treatment of MVM-infected cells with leptomycin B, a drug that specifically inhibits the CRM1-dependent nuclear export pathway, leads to a drastic accumulation of NS2 proteins in the nucleus. Both NS2 interaction with CRM1 and nuclear accumulation upon leptomycin B treatment strongly suggest that these nonstructural viral proteins are actively exported out of the nuclei of infected cells via a CRM1-mediated nuclear export pathway.
Figures





References
-
- Askjaer P, Jensen T H, Nilsson J, Englmeier L, Kjems J. The specificity of the CRM1-Rev nuclear export signal interaction is mediated by RanGTP. J Biol Chem. 1998;273:33414–33422. - PubMed
-
- Bartel P L, Chien C-T, Sternglanz R, Fields S. Using the two-hybrid system to detect protein-protein interactions. In: Hartley D A, editor. Cellular interactions in development: a practical approach. Oxford, United Kingdom: Oxford University Press; 1993. pp. 153–179.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
