

A role for RNA metabolism in inducing the heat shock response
- PMID: 10440229
- PMCID: PMC6174669
A role for RNA metabolism in inducing the heat shock response
Abstract
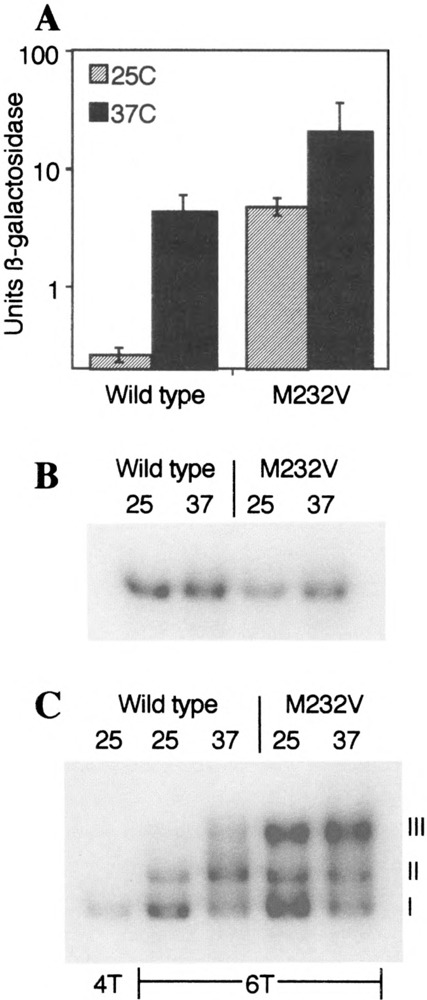
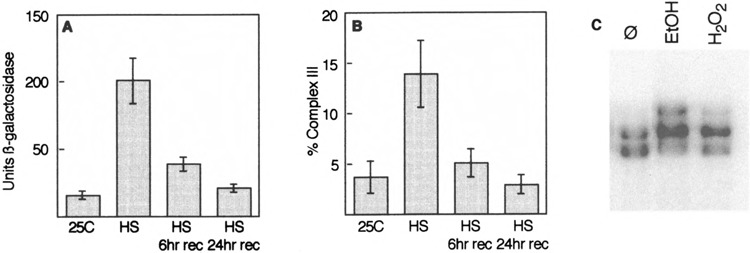
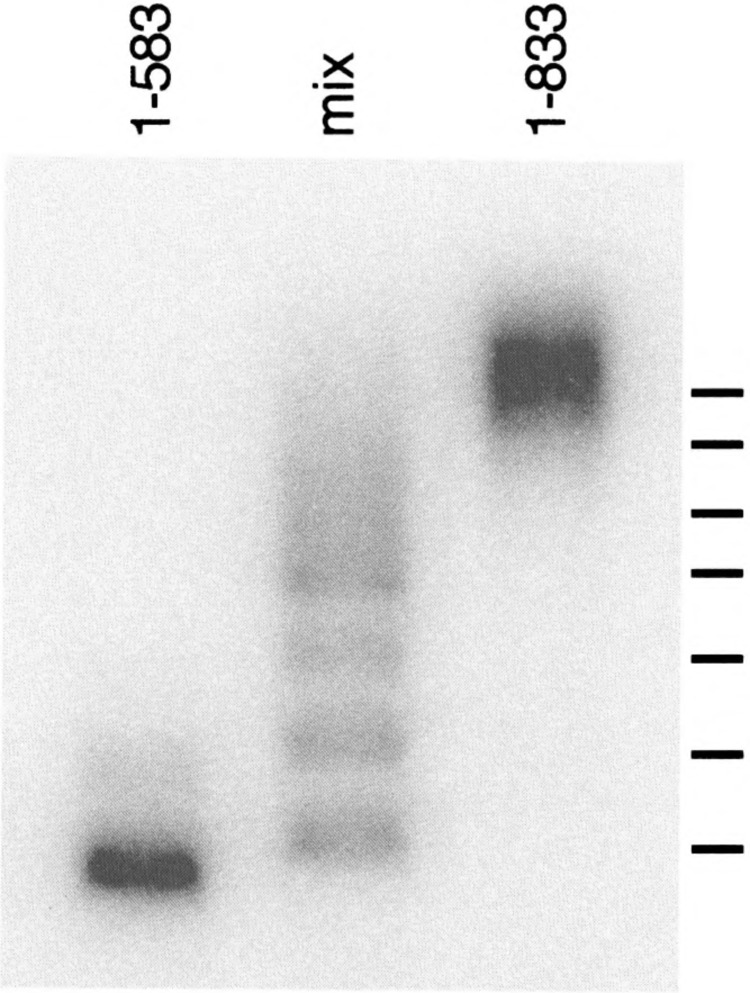
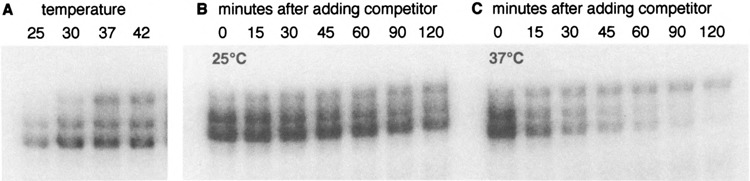
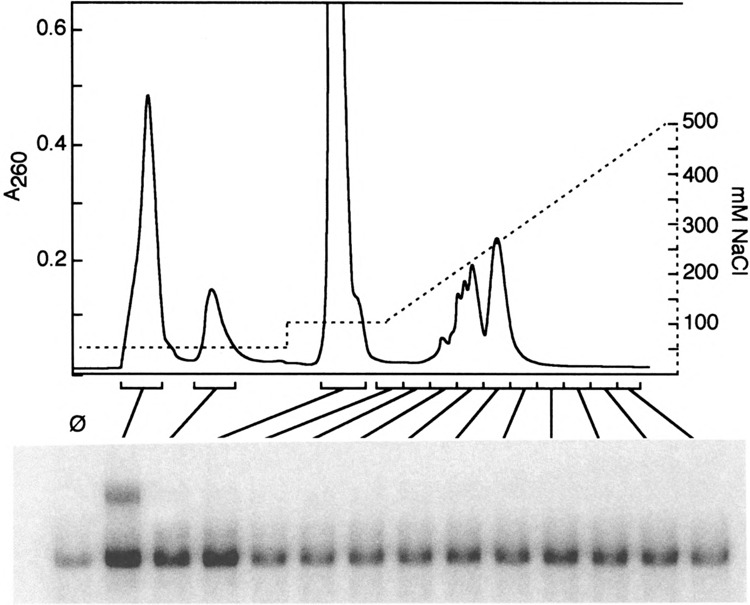
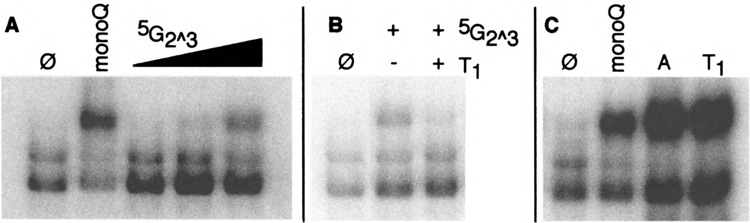
Yeast HSF is constitutively trimeric and DNA bound. Heat shock is thought to activate HSF by inducing a conformational change. We have developed an assay in which we can follow a conformational change of HSF that correlates with activity and thus appears to be the active conformation. This conformational change requires two HSF trimers bound cooperatively to DNA. The conformational change can be induced in whole cell extracts, and is thus amenable to biochemical analysis. We have purified a factor that triggers the conformational change. The factor is sensitive to dialysis, insensitive to NEM, and is not extractable by phenol. It is small, and apparently not a peptide. Mass spectroscopy identifies a novel guanine nucleotide that tracks with activity on columns. This novel nucleotide, purchased from Sigma, induces the conformational change (although this does not prove the identity of the activating factor unambiguously, because Sigma's preparation is contaminated with other compounds). What is the source of this nucleotide in cells? Activity can be generated by treating extracts with ribonuclease; this implicates RNA degradation as a source of HSF-activating activity. The heat shock response is primarily responsible for monitoring the levels of protein chaperones; how can RNA degradation be involved? Synthetic lethal interactions link HSF activity to ribosome biogenesis, suggesting a possible model. Ribosomal proteins are produced in large quantities, and in excess of rRNA; unassembled r-proteins are rapidly degraded (t1/2 approximately 3 min). Unassembled r-proteins aggregate readily. It is likely that unassembled r-proteins represent a major target of chaperones in vivo, and for proteasome-dependent degradation. Interference with rRNA processing (e.g., by heat shock) requires hsp70s to handle the aggregation-prone r-proteins, and proteasome proteins to help degrade the unassembled r-proteins before they aggregate. A nucleotide signal could be generated from the degradation products of the rRNA itself.
Figures
Similar articles
-
Complex regulation of the yeast heat shock transcription factor.Mol Biol Cell. 2000 May;11(5):1739-51. doi: 10.1091/mbc.11.5.1739. Mol Biol Cell. 2000. PMID: 10793148 Free PMC article.
-
Dynamic association of transcriptional activation domains and regulatory regions in Saccharomyces cerevisiae heat shock factor.Proc Natl Acad Sci U S A. 2002 Feb 5;99(3):1200-5. doi: 10.1073/pnas.032681299. Epub 2002 Jan 29. Proc Natl Acad Sci U S A. 2002. PMID: 11818569 Free PMC article.
-
A stress regulatory network for co-ordinated activation of proteasome expression mediated by yeast heat shock transcription factor.Mol Microbiol. 2006 Apr;60(1):240-51. doi: 10.1111/j.1365-2958.2006.05097.x. Mol Microbiol. 2006. PMID: 16556235
-
HSF transcription factor family, heat shock response, and protein intrinsic disorder.Curr Protein Pept Sci. 2012 Feb;13(1):86-103. doi: 10.2174/138920312799277956. Curr Protein Pept Sci. 2012. PMID: 22044151 Review.
-
[Advances in research of heat shock factor 1 and its relation with expression of heat shock protein].Zhonghua Shao Shang Za Zhi. 2012 Feb;28(1):78-80. Zhonghua Shao Shang Za Zhi. 2012. PMID: 22490544 Review. Chinese.
Cited by
-
The yeast heat shock transcription factor changes conformation in response to superoxide and temperature.Mol Biol Cell. 2000 May;11(5):1753-64. doi: 10.1091/mbc.11.5.1753. Mol Biol Cell. 2000. PMID: 10793149 Free PMC article.
References
-
- Abravaya K.; Myers M. P.; Murphy S. P.; Morimoto R. I. The human heat shock protein hsp70 interacts with HSF, the transcription factor that regulates heat shock gene expression. Genes Dev. 6:1153–1164; 1992. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases