

Genetic and biochemical evidence for the involvement of alpha-1,4 glucanotransferases in amylopectin synthesis
- PMID: 10444082
- PMCID: PMC59358
- DOI: 10.1104/pp.120.4.993
Genetic and biochemical evidence for the involvement of alpha-1,4 glucanotransferases in amylopectin synthesis
Abstract
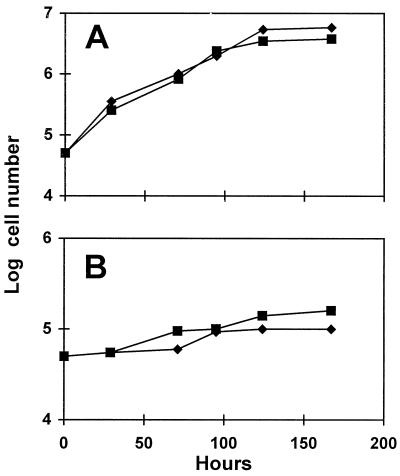
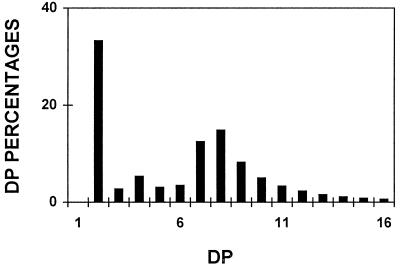
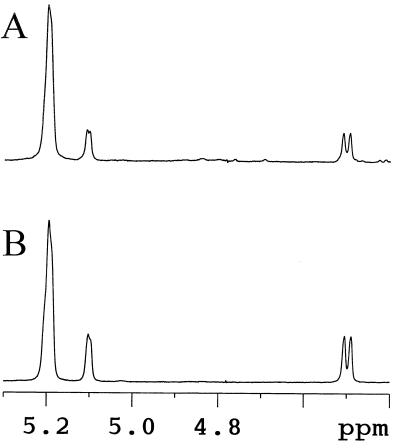
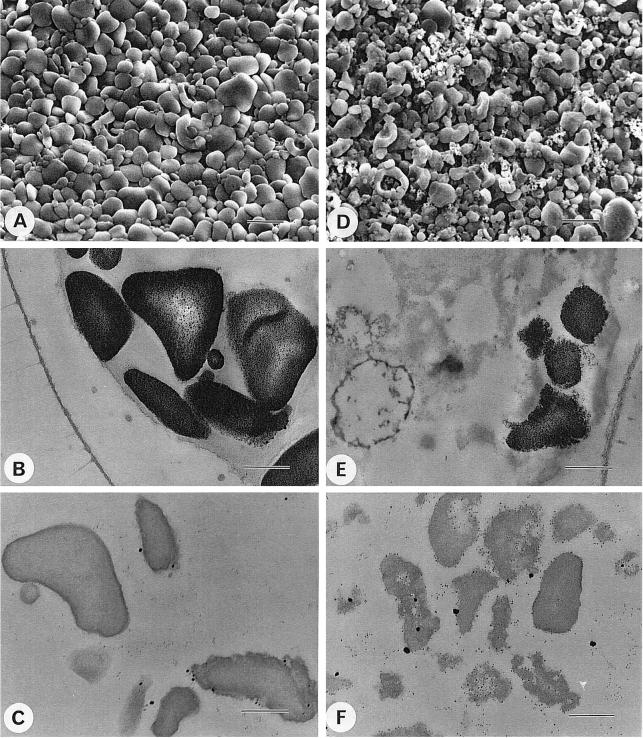
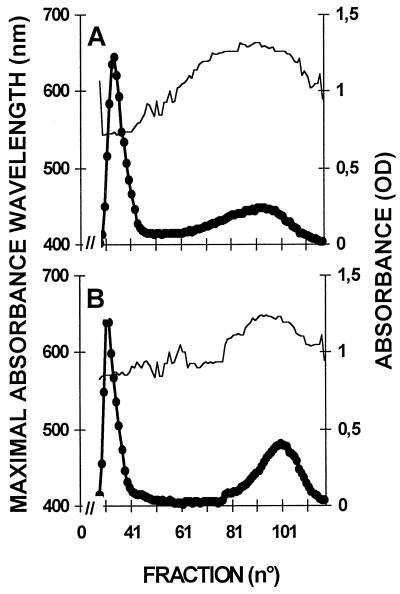
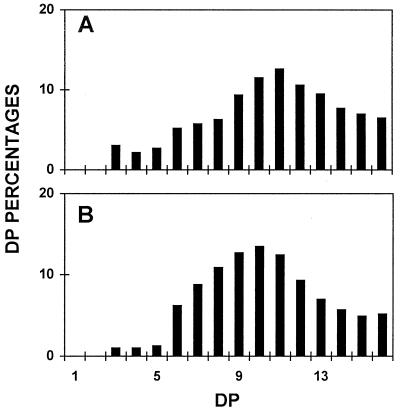
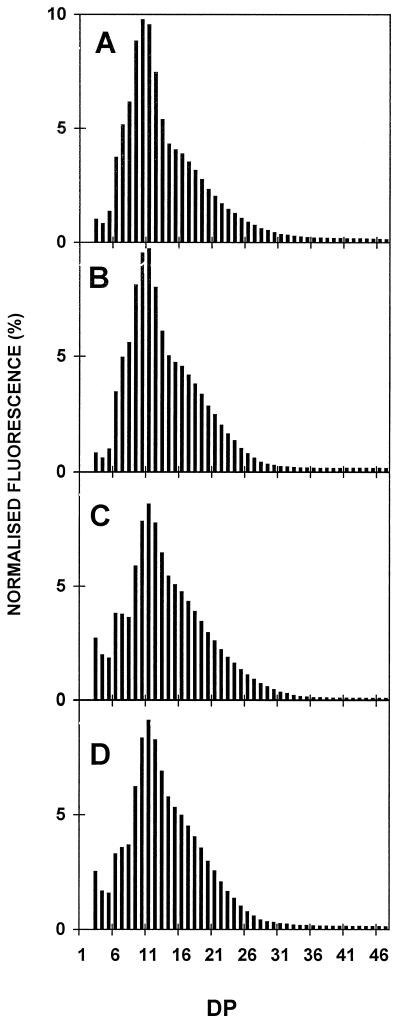
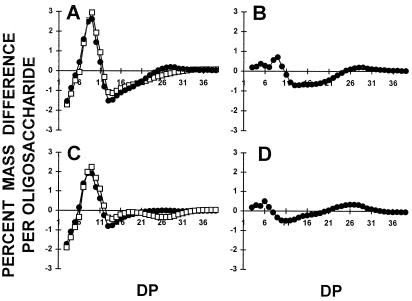
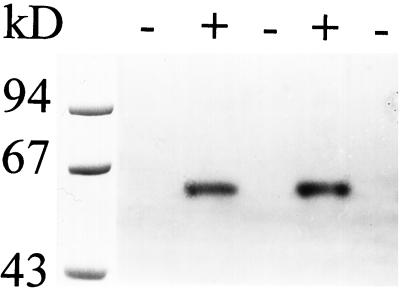
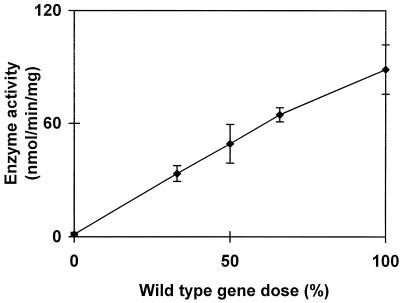
We describe a novel mutation in the Chlamydomonas reinhardtii STA11 gene, which results in significantly reduced granular starch deposition and major modifications in amylopectin structure and granule shape. This defect simultaneously leads to the accumulation of linear malto-oligosaccharides. The sta11-1 mutation causes the absence of an alpha-1,4 glucanotransferase known as disproportionating enzyme (D-enzyme). D-enzyme activity was found to be correlated with the amount of wild-type allele doses in gene dosage experiments. All other enzymes involved in starch biosynthesis, including ADP-glucose pyrophosphorylase, debranching enzymes, soluble and granule-bound starch synthases, branching enzymes, phosphorylases, alpha-glucosidases (maltases), and amylases, were unaffected by the mutation. These data indicate that the D-enzyme is required for normal starch granule biogenesis in the monocellular alga C. reinhardtii.
Figures

References
-
- Ball S. Regulation of starch biosynthesis. In: Rochaix J-D, Goldschmidt-Clermont M, Merchant S, Govindjee, editors. The Molecular Biology of Chloroplasts and Mitochondria in Chlamydomonas. Advances in Photosynthesis, Vol 7. Dordrecht, The Netherlands: Kluwer Academic Publishers; 1998. pp. 549–567.
-
- Ball S, Dirick L, Decq A, Martiat JC, Matagne RF. Plant Sci. 1990;66:1–9.
-
- Ball S, Guan H-P, James M, Myers A, Keeling P, Mouille G, Buléon A, Colonna P, Preiss J. From glycogen to amylopectin: a model explaining the biogenesis of the plant starch granule. Cell. 1996;86:349–352. - PubMed
-
- Ball S, Marianne T, Dirick L, Fresnoy M, Delrue B, Decq A. A Chlamydomonas reinhardtii low-starch mutant is defective for 3-phosphoglycerate activation and orthophosphate inhibition of ADP-glucose pyrophosphorylase. Planta. 1991;185:17–26. - PubMed
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
