

A cell-counting factor regulating structure size in Dictyostelium
- PMID: 10444594
- PMCID: PMC316923
- DOI: 10.1101/gad.13.15.1960
A cell-counting factor regulating structure size in Dictyostelium
Abstract
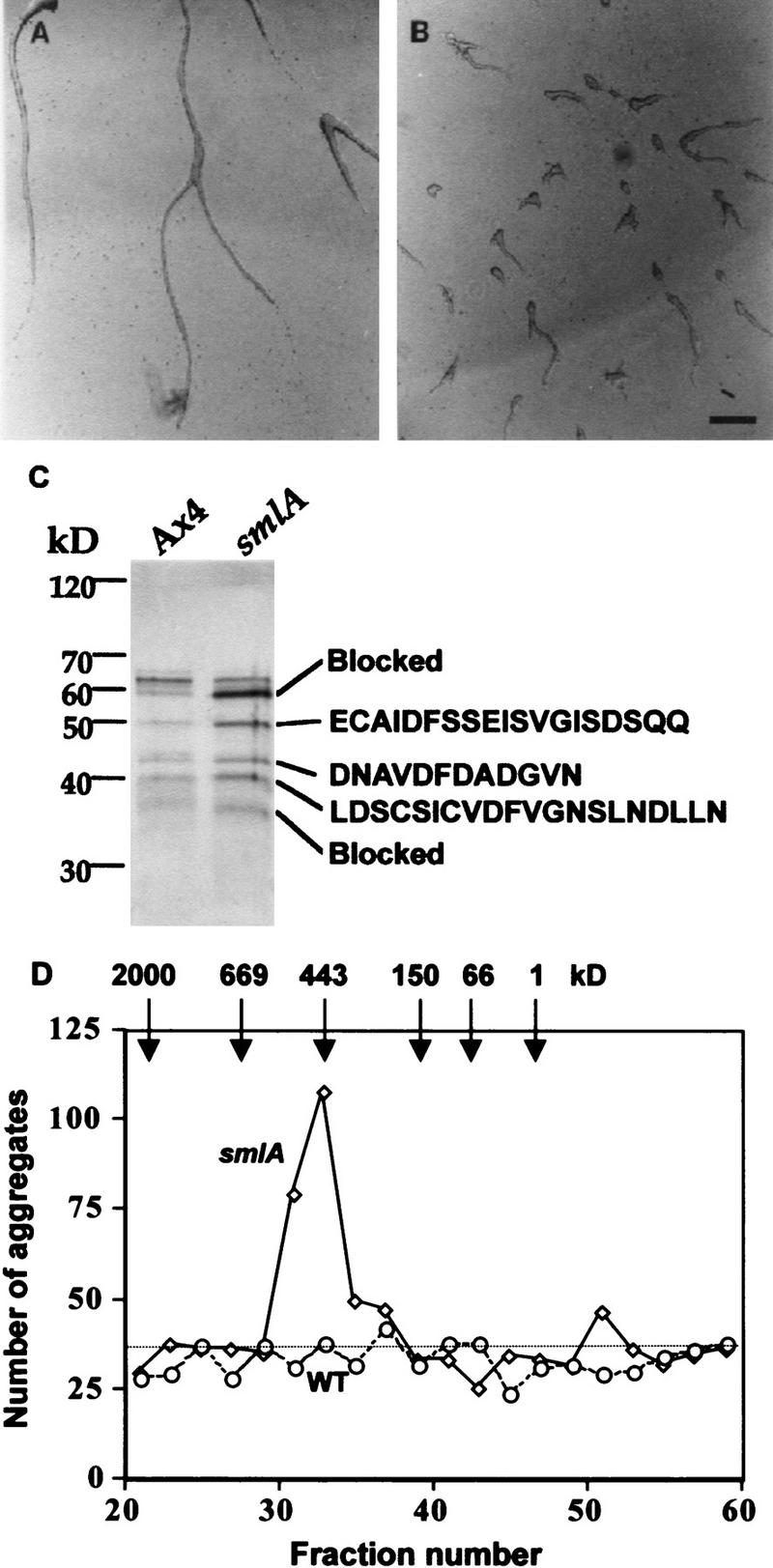
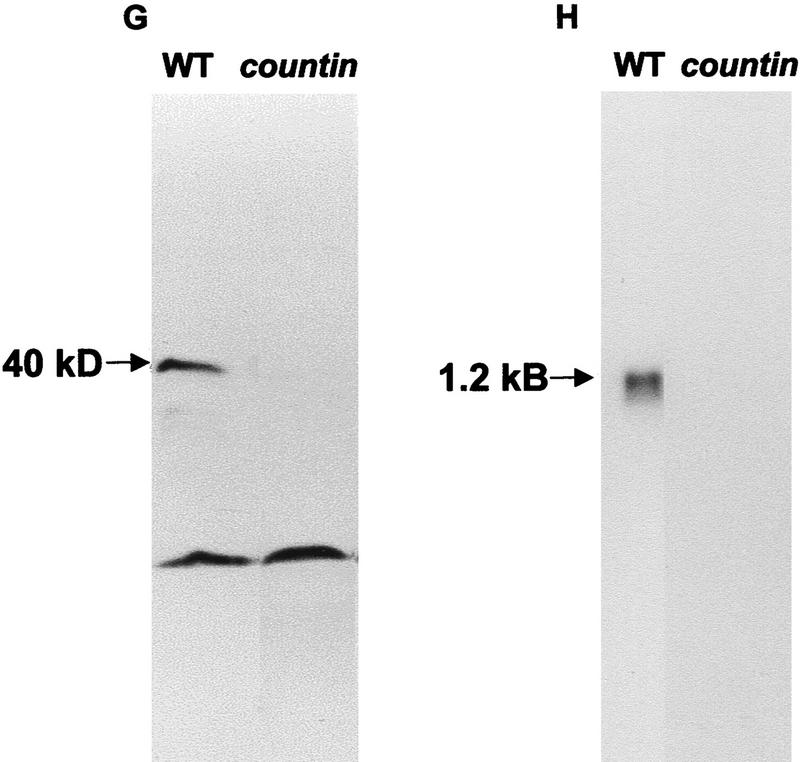
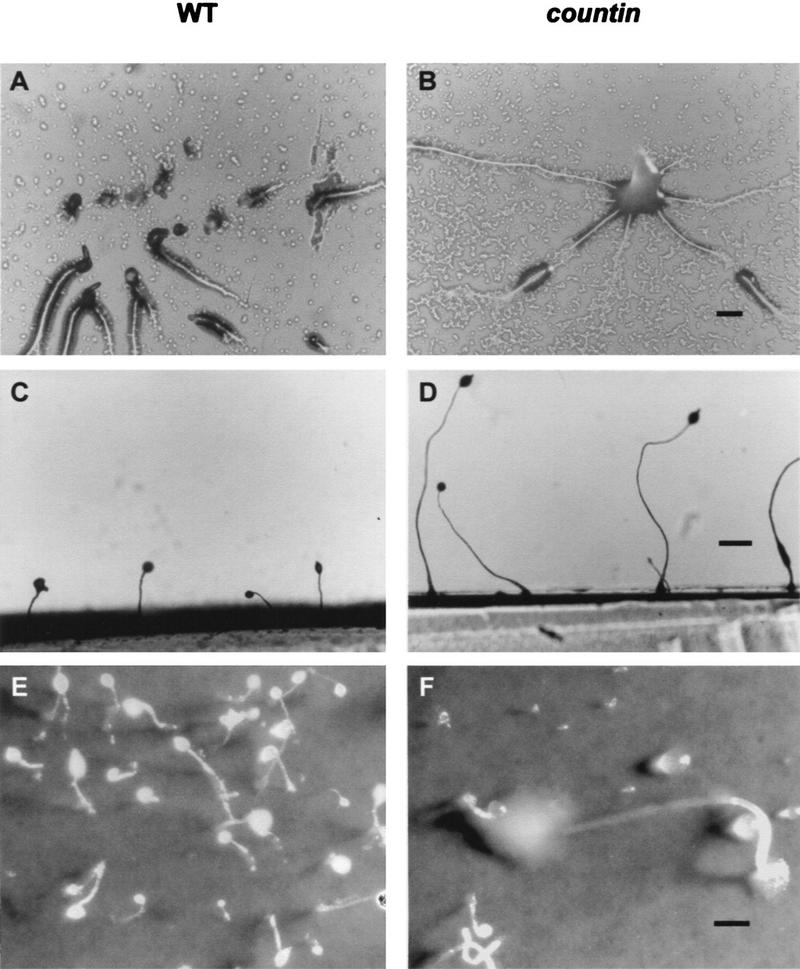
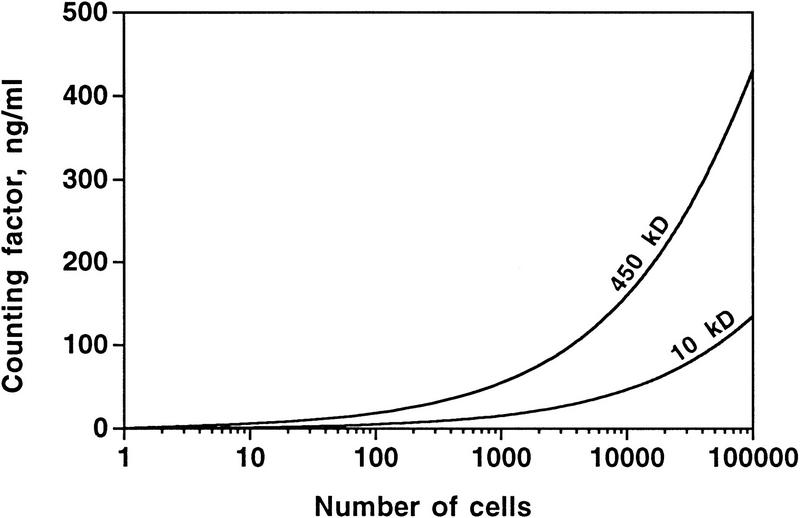
Developing Dictyostelium cells form large aggregation streams that break up into groups of 0.2 x 10(5) to 1 x 10(5) cells. Each group then becomes a fruiting body. smlA cells oversecrete an unknown factor that causes aggregation streams to break up into groups of approximately 5 x 10(3) cells and thus form very small fruiting bodies. We have purified the counting factor and find that it behaves as a complex of polypeptides with an effective molecular mass of 450 kD. One of the polypeptides is a 40-kD hydrophilic protein we have named counting. In transformants with a disrupted counting gene, there is no detectable secretion of counting factor, and the aggregation streams do not break up, resulting in huge (up to 2 x 10(5) cell) fruiting bodies.
Figures



References
-
- Alexander S. Developmental regulation and function of glycoproteins in Dictyostelium discoideum. In: Maeda Y, Inouye K, Takeuchi I, editors. Dictyostelium - A model system for cell and developmental biology. Tokyo, Japan: Universal Academy Press; 1997. pp. 349–362.
-
- Bonner JT, Hoffman ME. Evidence for a substance responsible for spacing pattern of aggregation and fruiting bodies in the cellular slime mold. J Embryol Exp Morphol. 1963;11:571–589. - PubMed
-
- Brock DA, Buczynski F, Spann TP, Wood SA, Cardelli J, Gomer RH. A Dictyostelium mutant with defective aggregate size determination. Development. 1996;122:2569–2578. - PubMed
-
- Clarke M, Gomer RH. PSF and CMF, autocrine factors that regulate gene expression during growth and early development of Dictyostelium. Experientia. 1995;51:1124–1134. - PubMed
-
- Compton T. Degenerate primers for DNA amplification. In: Innis MA, Gelfand DH, Sninsky JJ, White TJ, editors. PCR protocols. A guide to methods and applications. San Diego, CA: Academic Press; 1990. pp. 39–45.
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials