

Administration of interleukin-12 exerts a therapeutic instead of a long-term preventive effect on mite Der p I allergen-induced animal model of airway inflammation
- PMID: 10447737
- PMCID: PMC2326828
- DOI: 10.1046/j.1365-2567.1999.00768.x
Administration of interleukin-12 exerts a therapeutic instead of a long-term preventive effect on mite Der p I allergen-induced animal model of airway inflammation
Abstract
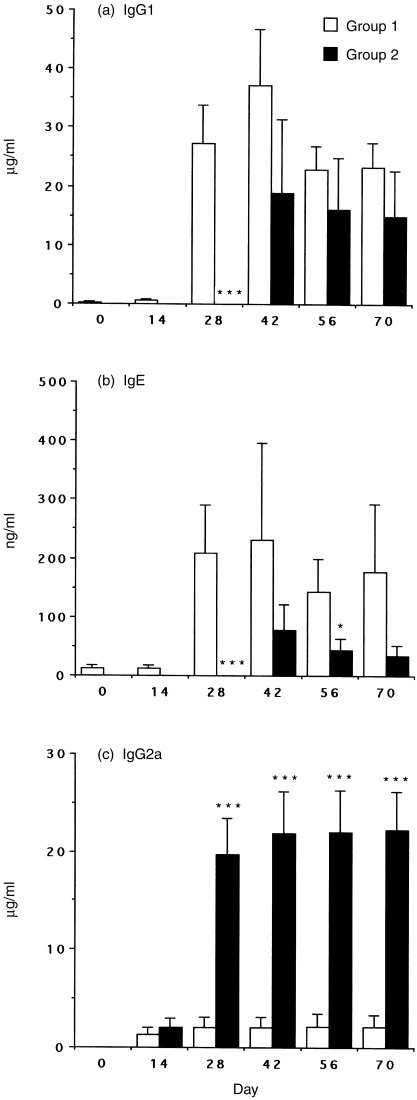
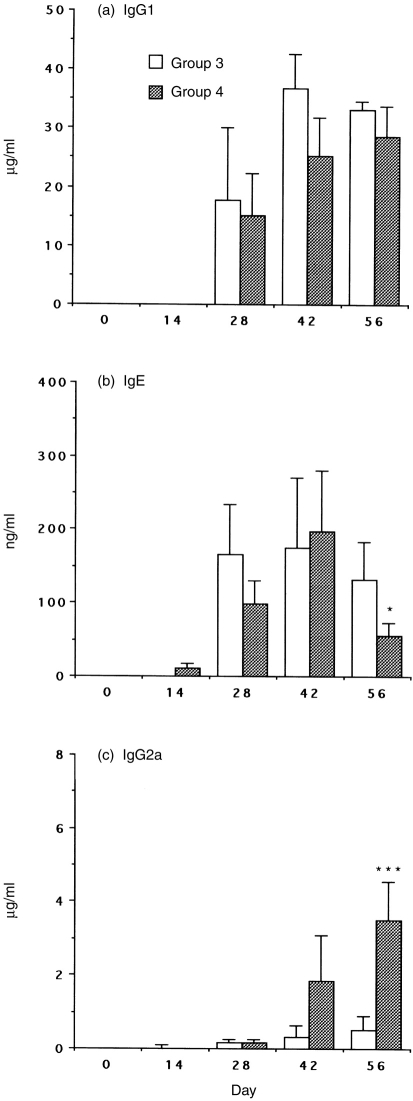
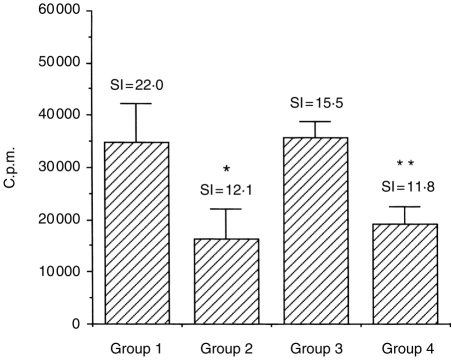
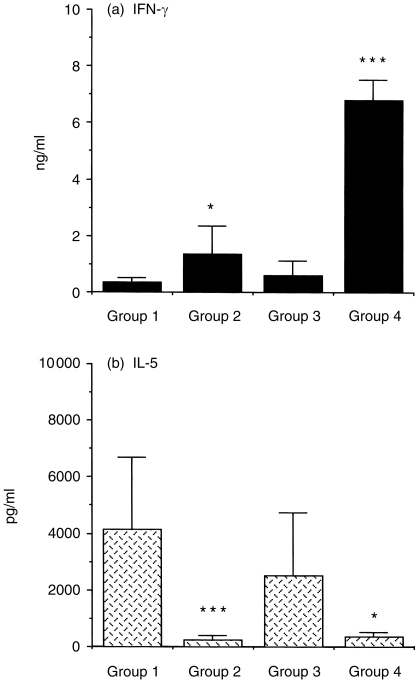
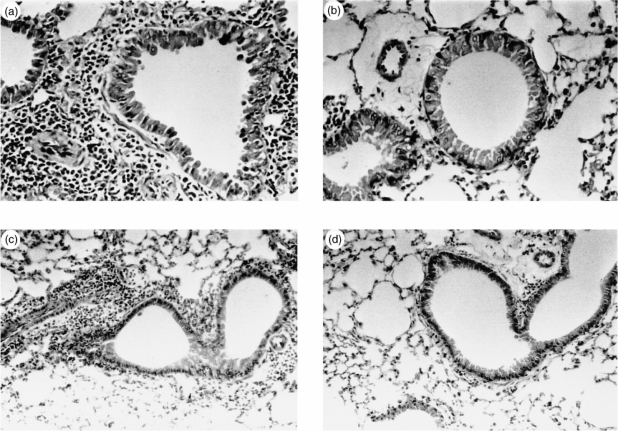
Interleukin-12 (IL-12) is a key cytokine, which promotes T helper type 1 (Th1) cell-mediated immunity and inhibits Th2-type responses. It has been previously shown that IL-12 administration during active immunization following a single allergen exposure can prevent antigen-induced increases in immunoglobulin E (IgE) formation, Th2 cytokine production and bronchoalveolar lavage (BAL) eosinophils in a murine model of allergic airway inflammation. Thus, these studies have now been extended and two IL-12 treatment protocols on this murine model were evaluated. Administration of IL-12 during the active immunization strikingly increased Der p I-specific serum IgG2a and transiently decreased the levels of IgG1 and IgE antibodies following multiple allergen challenges. Such early treatment of IL-12 down-regulated IL-5 production and modestly up-regulated interferon-gamma production but did not effect BAL eosinophilia. These results suggest that repeated exposure to antigen and IL-12 is necessary to maintain a persistent Th1-recall response. Furthermore, administration of IL-12 to actively immunized mice, in which Th2-associated responses were established, had a significant effect on IgG2a synthesis and a modest effect on IgE levels, also down-regulation of IL-5 production, and markedly increased interferon-gamma production and abolished recruitment of eosinophils. Therefore, these data indicate that IL-12 can inhibit antigen-induced eosinophil infiltration into airways, despite the existence of a Th2-associated response. Taken together, these studies suggest that IL-12 may be useful as an immunotherapeutic agent in the treatment of such pulmonary allergic disorders as bronchial asthma.
Figures

Similar articles
-
Administration of interleukin-12 prevents mite Der p 1 allergen-IgE antibody production and airway eosinophil infiltration in an animal model of airway inflammation.Scand J Immunol. 1999 Mar;49(3):229-36. doi: 10.1046/j.1365-3083.1999.00503.x. Scand J Immunol. 1999. PMID: 10102639
-
Absence of immunoglobulin E synthesis and airway eosinophilia by vaccination with plasmid DNA encoding ProDer p 1.Clin Exp Allergy. 2003 Feb;33(2):218-25. doi: 10.1046/j.1365-2222.2003.01584.x. Clin Exp Allergy. 2003. PMID: 12580915
-
Interleukin 12 inhibits antigen-induced airway hyperresponsiveness, inflammation, and Th2 cytokine expression in mice.J Exp Med. 1995 Nov 1;182(5):1527-36. doi: 10.1084/jem.182.5.1527. J Exp Med. 1995. PMID: 7595222 Free PMC article.
-
Immunologic responses following respiratory sensitization to house dust mite allergens in mice.Immunol Cell Biol. 1996 Apr;74(2):174-9. doi: 10.1038/icb.1996.24. Immunol Cell Biol. 1996. PMID: 8724006 Review.
-
Biological properties of interleukin-12 and its therapeutic use in persistent hepatitis B virus and hepatitis C virus infection.J Viral Hepat. 1997;4 Suppl 2:83-6. doi: 10.1111/j.1365-2893.1997.tb00184.x. J Viral Hepat. 1997. PMID: 9429214 Review.
Cited by
-
Anti-inflammatory cytokines in asthma and allergy: interleukin-10, interleukin-12, interferon-gamma.Mediators Inflamm. 2001 Apr;10(2):51-9. doi: 10.1080/09629350120054518. Mediators Inflamm. 2001. PMID: 11405550 Free PMC article. Review.
-
Maternal allergy increases susceptibility to offspring allergy in association with TH2-biased epigenetic alterations in a mouse model of peanut allergy.J Allergy Clin Immunol. 2014 Dec;134(6):1339-1345.e7. doi: 10.1016/j.jaci.2014.08.034. Epub 2014 Oct 16. J Allergy Clin Immunol. 2014. PMID: 25441650 Free PMC article.
-
Administration of polysaccharides from Antrodia camphorata modulates dendritic cell function and alleviates allergen-induced T helper type 2 responses in a mouse model of asthma.Immunology. 2010 Mar;129(3):351-62. doi: 10.1111/j.1365-2567.2009.03175.x. Epub 2009 Nov 11. Immunology. 2010. PMID: 19909376 Free PMC article.
-
Asthma and therapeutics: recombinant therapies in asthma.Allergy Asthma Clin Immunol. 2005 Mar 15;1(1):34-41. doi: 10.1186/1710-1492-1-1-34. Epub 2005 Mar 15. Allergy Asthma Clin Immunol. 2005. PMID: 20529233 Free PMC article.
-
Serotonin decreases alveolar epithelial fluid transport via a direct inhibition of the epithelial sodium channel.Am J Respir Cell Mol Biol. 2010 Jul;43(1):99-108. doi: 10.1165/rcmb.2008-0472OC. Epub 2009 Aug 28. Am J Respir Cell Mol Biol. 2010. PMID: 19717814 Free PMC article.
References
-
- Voorhorst R, Spieksma-Boezeman MIA, Spieksma FTM. Is a mite (Dermatophagoides sp) the producer of the house dust allergen? Allergy Asthma. 1964;10:329. - PubMed
-
- Krillis S, Baldo BA, Basten A. Antigens and allergens from the common house dust mite Dermatophagoides pteronyssinus. Part II. Identification of the major IgE binding antigen by crossed radioimmunoelectrophoresis. J Allergy Clin Immunol. 1984;74:142. - PubMed
-
- Platts-Mills TA, Snajdr MJ, Ishizaka K, Frankland AW. Measurement of IgE antibody by an antigen-binding assay: correlation with PK activity and IgG and IgA antibodies to allergens. J Immunol. 1978;120:1201. - PubMed
-
- Desreumaux P, Capron M. Eosinophils in allergic reactions. Curr Opin Immunol. 1996;8:790. - PubMed
-
- Busse WW, Calhoun WF, Sedgwick JD. Mechanism of airway inflammation in asthma. Am Rev Respir Dis. 1993;147:20. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
