

Interactions of TLC1 (which encodes the RNA subunit of telomerase), TEL1, and MEC1 in regulating telomere length in the yeast Saccharomyces cerevisiae
- PMID: 10454554
- PMCID: PMC84515
- DOI: 10.1128/MCB.19.9.6065
Interactions of TLC1 (which encodes the RNA subunit of telomerase), TEL1, and MEC1 in regulating telomere length in the yeast Saccharomyces cerevisiae
Abstract
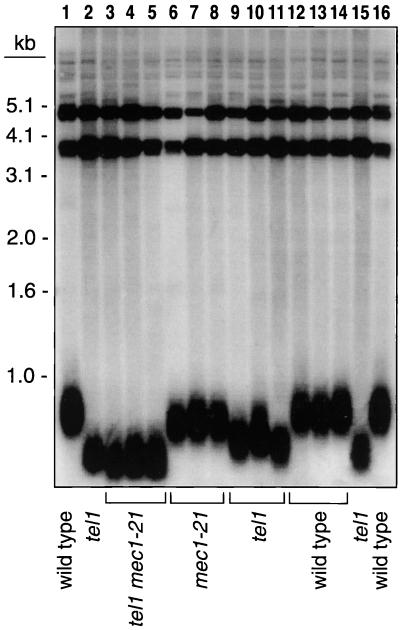
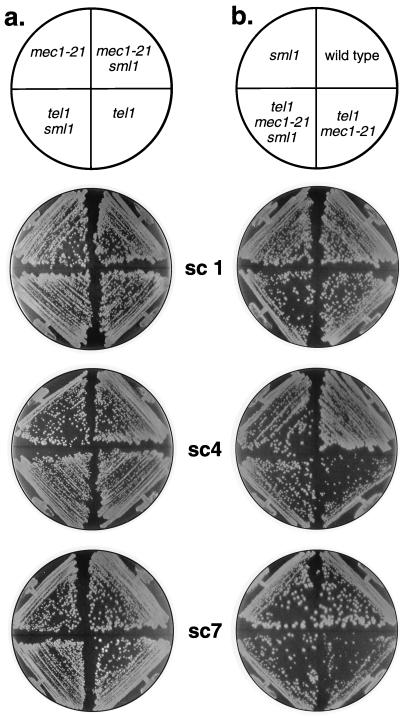
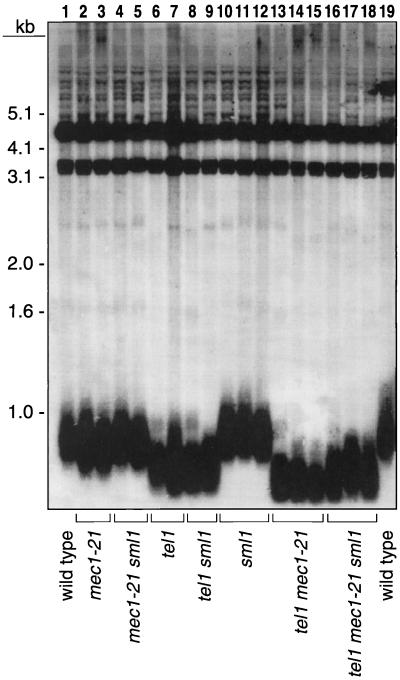
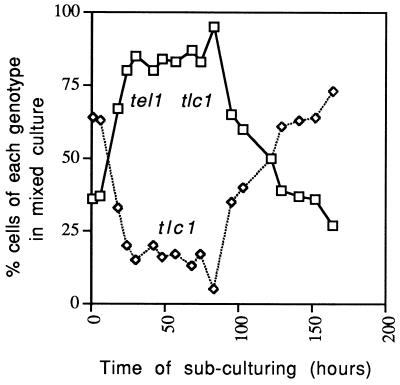
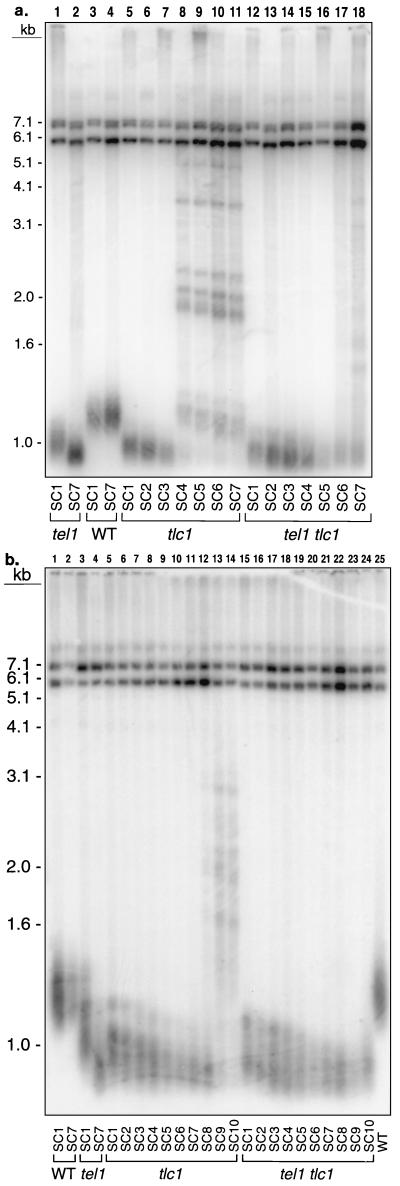
In the yeast Saccharomyces cerevisiae, chromosomes terminate with a repetitive sequence [poly(TG(1-3))] 350 to 500 bp in length. Strains with a mutation of TEL1, a homolog of the human gene (ATM) mutated in patients with ataxia telangiectasia, have short but stable telomeric repeats. Mutations of TLC1 (encoding the RNA subunit of telomerase) result in strains that have continually shortening telomeres and a gradual loss of cell viability; survivors of senescence arise as a consequence of a Rad52p-dependent recombination events that amplify telomeric and subtelomeric repeats. We show that a mutation in MEC1 (a gene related in sequence to TEL1 and ATM) reduces telomere length and that tel1 mec1 double mutant strains have a senescent phenotype similar to that found in tlc1 strains. As observed in tlc1 strains, survivors of senescence in the tel1 mec1 strains occur by a Rad52p-dependent amplification of telomeric and subtelomeric repeats. In addition, we find that strains with both tel1 and tlc1 mutations have a delayed loss of cell viability compared to strains with the single tlc1 mutation. This result argues that the role of Tel1p in telomere maintenance is not solely a direct activation of telomerase.
Figures




References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous