

beta-Chemokines enhance parasite uptake and promote nitric oxide-dependent microbiostatic activity in murine inflammatory macrophages infected with Trypanosoma cruzi
- PMID: 10456936
- PMCID: PMC96814
- DOI: 10.1128/IAI.67.9.4819-4826.1999
beta-Chemokines enhance parasite uptake and promote nitric oxide-dependent microbiostatic activity in murine inflammatory macrophages infected with Trypanosoma cruzi
Abstract
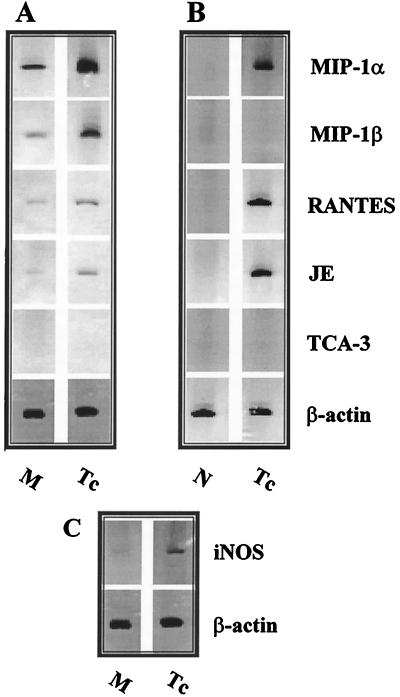
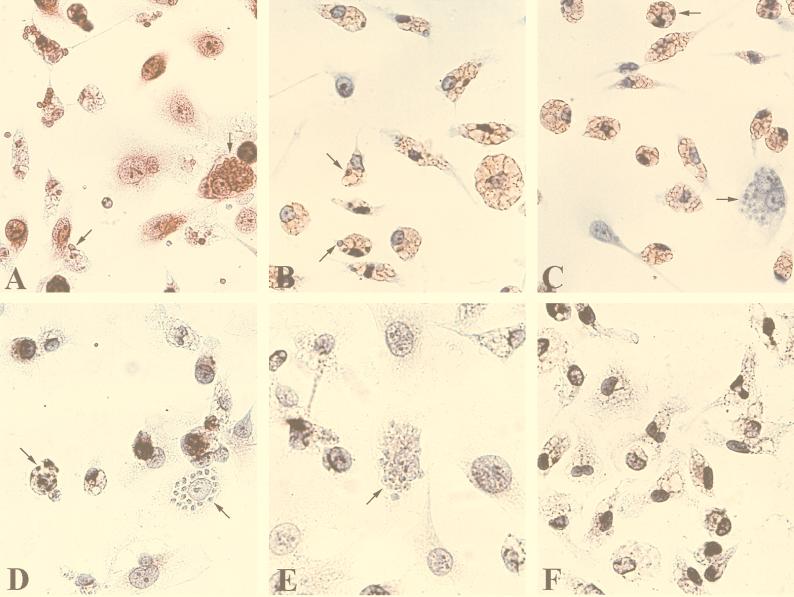
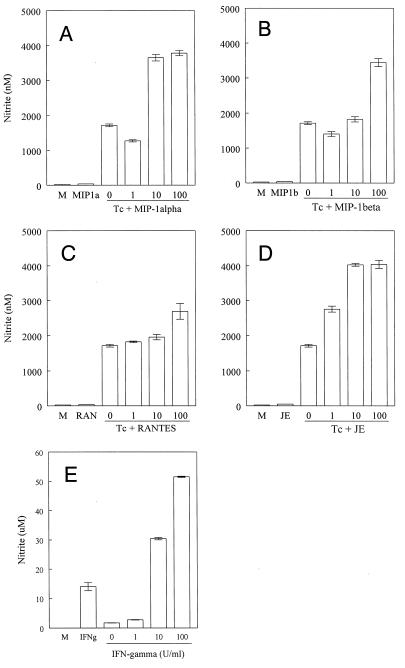
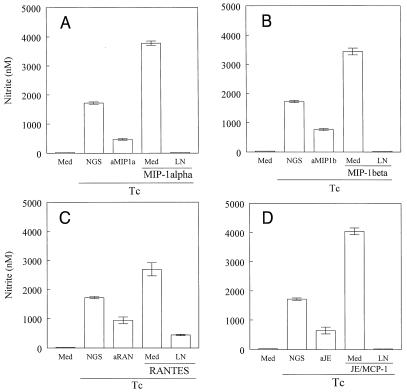
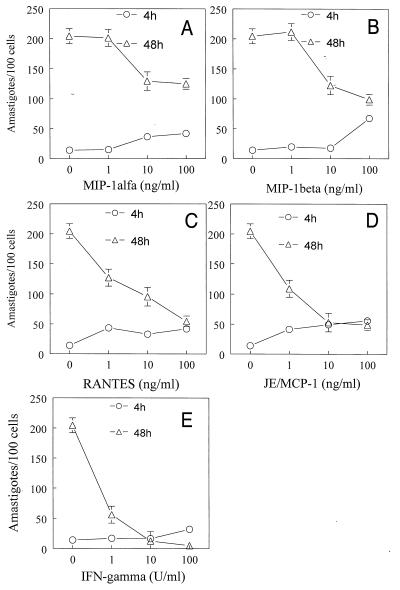
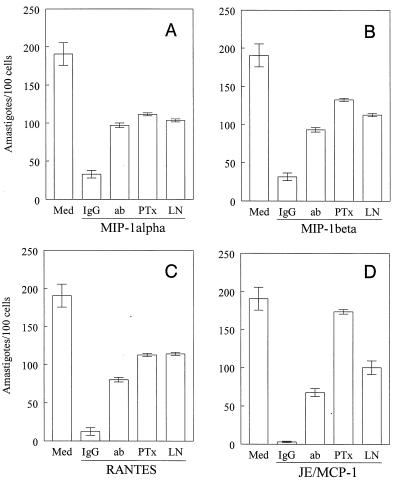
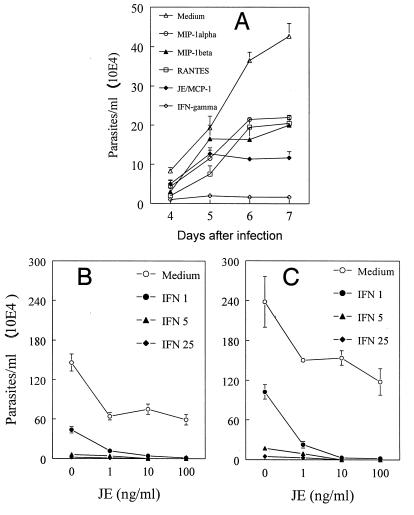
In the present study, we describe the ability of Trypanosoma cruzi trypomastigotes to stimulate the synthesis of beta-chemokines by macrophages. In vivo infection with T. cruzi led to MIP-1alpha, RANTES, and JE/MCP1 mRNA expression by cells from peritoneal inflammatory exudate. In addition, in vitro infection with T. cruzi resulted in expression of beta-chemokine MIP-1alpha, MIP-1beta, RANTES, and JE mRNA by macrophages. The expression of the beta-chemokine MIP-1alpha, MIP-1beta, RANTES, and JE proteins by murine macrophages cultured with trypomastigote forms of T. cruzi was confirmed by immunocytochemistry. Interestingly, macrophage infection with T. cruzi also resulted in NO production, which we found to be mediated mainly by beta-chemokines. Hence, treatment with anti-beta-chemokine-specific neutralizing antibodies partially inhibited NO release by macrophages incubated with T. cruzi parasites. Further, the addition of the exogenous beta-chemokines MIP-1alpha, MIP-1beta, RANTES, and JE/MCP-1 induced an increased T. cruzi uptake, leading to enhanced NO production and control of parasite replication in a dose-dependent manner. L-NMMA, a specific inhibitor of the L-arginine-NO pathway, caused a decrease in NO production and parasite killing when added to cultures of macrophages stimulated with beta-chemokines. Among the beta-chemokines tested, JE was more potent in inhibiting parasite growth, although it was much less efficient than gamma interferon (IFN-gamma). Nevertheless, JE potentiates parasite killing by macrophages incubated with low doses of IFN-gamma. Together, these results suggest that in addition to their chemotactic activity, murine beta-chemokines may also contribute to enhancing parasite uptake and promoting control of parasite replication in macrophages and may play a role in resistance to T. cruzi infection.
Figures
References
-
- Andrade Z A. Mechanisms of myocardial damage in Trypanosoma cruzi infection. Ciba Found Symp. 1983;99:214–233. - PubMed
-
- Baggiolini M, Dewald B, Moser B. Interleukin-8 and related chemotactic cytokines—CXC and CC chemokines. Adv Immunol. 1994;55:97–179. - PubMed
-
- Brener Z, Gazzinelli R T. Immunological control of Trypanosoma cruzi infection and pathogenesis of Chagas’ disease. Int Arch Allergy Immunol. 1997;114:103–110. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
