

Signalling of static and dynamic features of muscle spindle input by external cuneate neurones in the cat
- PMID: 10457071
- PMCID: PMC2269501
- DOI: 10.1111/j.1469-7793.1999.0559m.x
Signalling of static and dynamic features of muscle spindle input by external cuneate neurones in the cat
Abstract
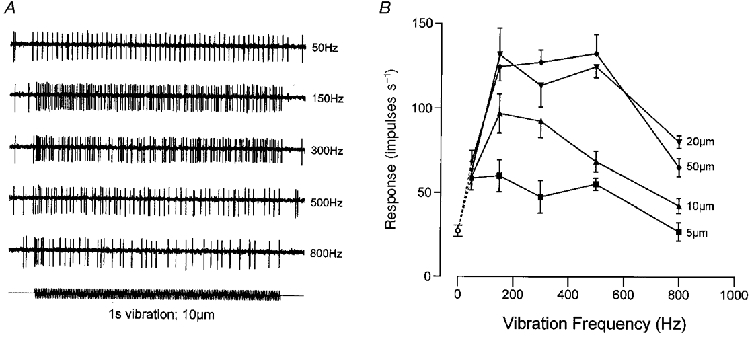
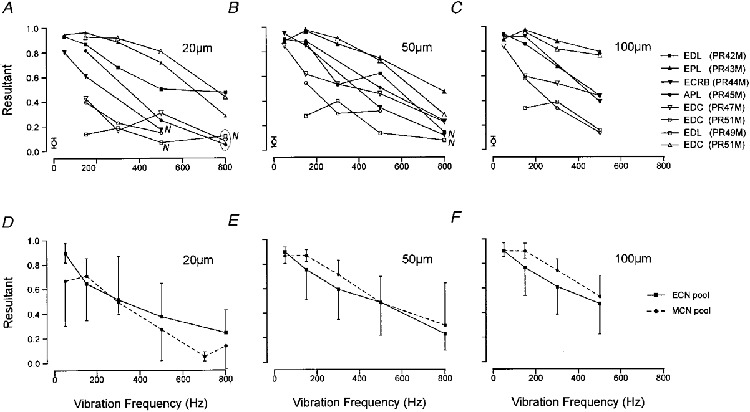
1. The present experiments examined the capacity of external cuneate nucleus (ECN) neurones in the anaesthetized cat to respond to static and vibrotactile stretch of forearm extensor muscles. The aim was to compare their signalling capacities with the known properties of main cuneate neurones in order to determine whether there is differential processing of muscle spindle inputs at these parallel relay sites. 2. Static stretch (<= 2 mm in amplitude) and sinusoidal vibration were applied longitudinally to individual muscle tendons and responses recorded from single ECN neurones. The muscle-related ECN neurones that were sampled displayed a high sensitivity to both static and dynamic components of stretch, including muscle vibration at frequencies of 50-800 Hz, consistent with their dominant input being derived from primary spindle afferent fibres. 3. In response to ramp-and-hold muscle stretch, ECN neurones resembled their main cuneate counterparts in the pattern of their responses and in quantitative response measures. Their coefficients of variation in interspike intervals during steady stretch ranged from approximately 0.3 to 0.7, as they do in main cuneate responses, and their stimulus-response relations were graded as a function of stretch magnitude with low variability in responses at a fixed stretch amplitude. 4. In response to muscle vibration, ECN activity was tightly phase locked to the vibration waveform, in particular at frequencies of <= 150 Hz, where vector strength measures (R) were high (R >= 0.8) before declining as a function of frequency, with R values of approximately 0.6 at 300 Hz and <= 0.4 at 800 Hz. Both the qualitative and quantitative aspects of ECN responsiveness to the vibro-stretch disturbances were indistinguishable from those of the main cuneate neurones. 5. The results demonstrate a high transmission fidelity for muscle signals across the ECN and no evidence for differential synaptic transmission across the parallel main and external cuneate nuclei. Earlier limitations observed in the capacity of cerebellar Purkinje cells to respond to primary spindle inputs must therefore be imposed at synapses within the cerebellum.
Figures


Similar articles
-
Signalling of static and dynamic features of muscle spindle input by cuneate neurones in the cat.J Physiol. 1998 Aug 1;510 ( Pt 3)(Pt 3):923-39. doi: 10.1111/j.1469-7793.1998.923bj.x. J Physiol. 1998. PMID: 9660903 Free PMC article.
-
Coding of information about tactile stimuli by neurones of the cuneate nucleus.J Physiol. 1978 Dec;285:493-513. doi: 10.1113/jphysiol.1978.sp012585. J Physiol. 1978. PMID: 745115 Free PMC article.
-
The effects of neonatal median nerve injury on the responsiveness of tactile neurones within the cuneate nucleus of the cat.J Physiol. 1997 Dec 15;505 ( Pt 3)(Pt 3):759-68. doi: 10.1111/j.1469-7793.1997.759ba.x. J Physiol. 1997. PMID: 9457650 Free PMC article.
-
Temporal patterning in the responses of gracile and cuneate neurones in the cat to cutaneous vibration.J Physiol. 1987 May;386:277-91. doi: 10.1113/jphysiol.1987.sp016534. J Physiol. 1987. PMID: 3681710 Free PMC article.
-
Responses of neurones in motor cortex and in area 3A to controlled stretches of forelimb muscles in cebus monkeys.J Physiol. 1975 Oct;251(3):833-53. doi: 10.1113/jphysiol.1975.sp011125. J Physiol. 1975. PMID: 127038 Free PMC article.
Cited by
-
Diverse precerebellar neurons share similar intrinsic excitability.J Neurosci. 2011 Nov 16;31(46):16665-74. doi: 10.1523/JNEUROSCI.3314-11.2011. J Neurosci. 2011. PMID: 22090493 Free PMC article.
-
Processing afferent proprioceptive information at the main cuneate nucleus of anesthetized cats.J Neurosci. 2010 Nov 17;30(46):15383-99. doi: 10.1523/JNEUROSCI.2193-10.2010. J Neurosci. 2010. PMID: 21084595 Free PMC article.
-
Gating of reafference in the external cuneate nucleus during self-generated movements in wake but not sleep.Elife. 2016 Aug 3;5:e18749. doi: 10.7554/eLife.18749. Elife. 2016. PMID: 27487470 Free PMC article.
-
Facilitation of mossy fibre-driven spiking in the cerebellar nuclei by the synchrony of inhibition.J Physiol. 2017 Aug 1;595(15):5245-5264. doi: 10.1113/JP274321. Epub 2017 Jun 11. J Physiol. 2017. PMID: 28513836 Free PMC article.
-
Proprioception: A New Era Set in Motion by Emerging Genetic and Bionic Strategies?Annu Rev Physiol. 2023 Feb 10;85:1-24. doi: 10.1146/annurev-physiol-040122-081302. Epub 2022 Nov 18. Annu Rev Physiol. 2023. PMID: 36400128 Free PMC article. Review.
References
-
- Andersen P, Eccles JC, Oshima T, Schmidt RF. Mechanisms of synaptic transmission in the cuneate nucleus. Journal of Neurophysiology. 1964;27:1096–1116. - PubMed
-
- Barker D. The innervation of mammalian skeletal muscle. In: De Reuck AVS, Knight J, editors. Myotatic, Kinesthetic and Vestibular Mechanisms, Ciba Foundation Symposium. London: Churchill; 1967. pp. 3–15.
-
- Berman AL. The Brain Stem of the Cat; A Cytoarchitectonic Atlas with Stereotaxic Coordinates. Madison: University of Wisconsin Press; 1968.
-
- Bianconi R, Van Der Meulen JP. The response to vibration of the end organs of the mammalian muscle spindles. Journal of Neurophysiology. 1963;26:177–190. - PubMed
-
- Bledsoe SC, Rupert AL, Moushegian G. Response characteristics of cochlear nucleus neurons to 500 Hz tones and noise: findings relating to frequency-following potentials. Journal of Neurophysiology. 1982;47:113–127. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous