

Control of apical membrane chloride permeability in the renal A6 cell line by nucleotides
- PMID: 10457087
- PMCID: PMC2269550
- DOI: 10.1111/j.1469-7793.1999.0737n.x
Control of apical membrane chloride permeability in the renal A6 cell line by nucleotides
Abstract
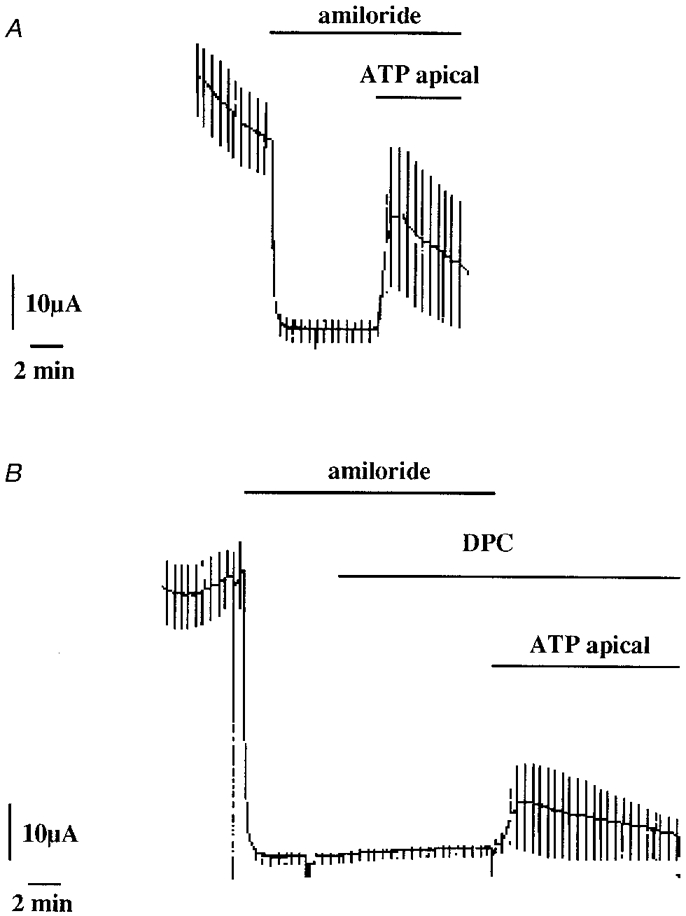
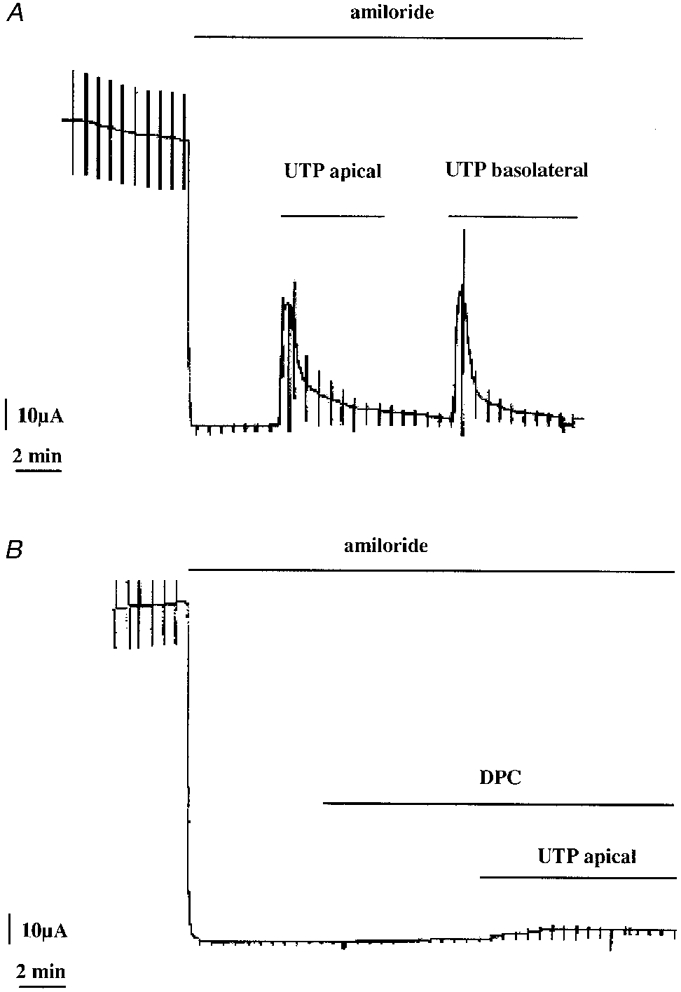
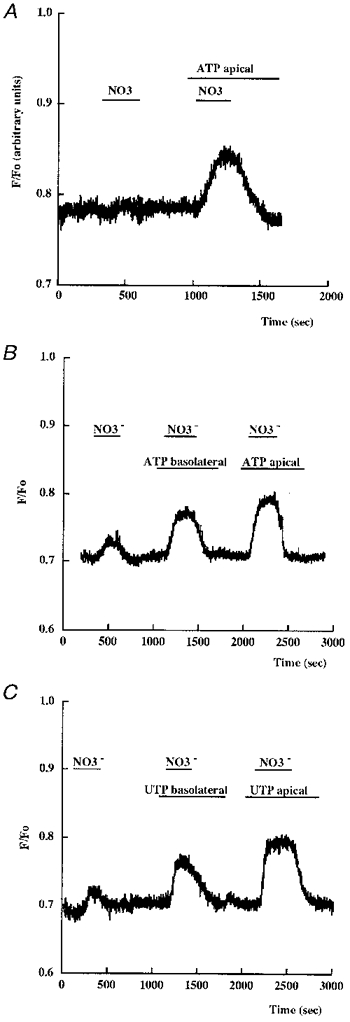
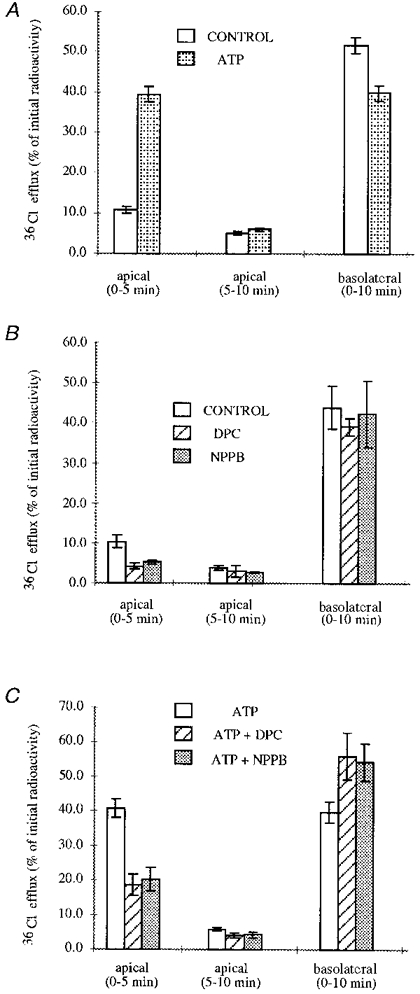
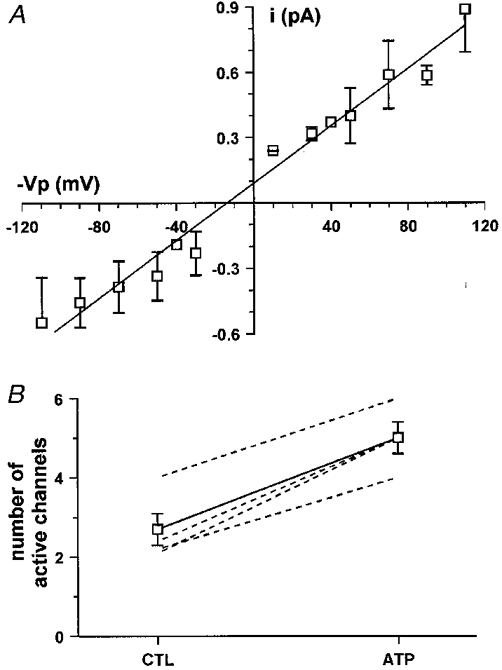
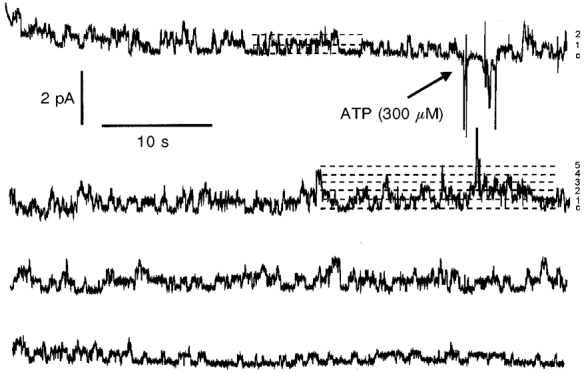
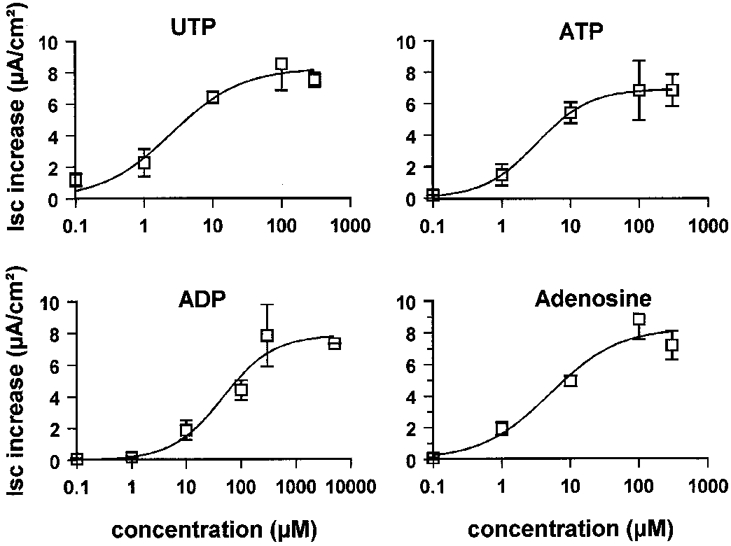
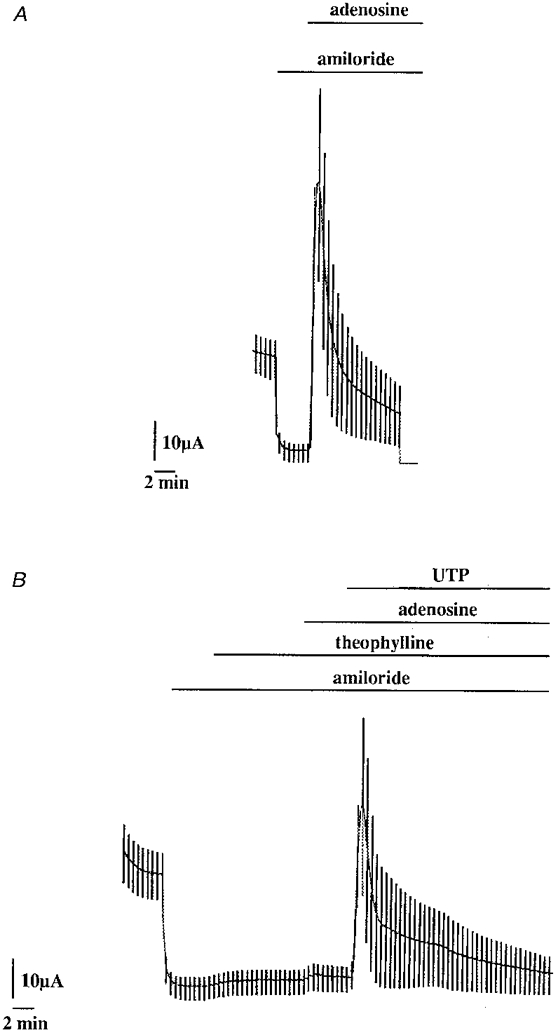
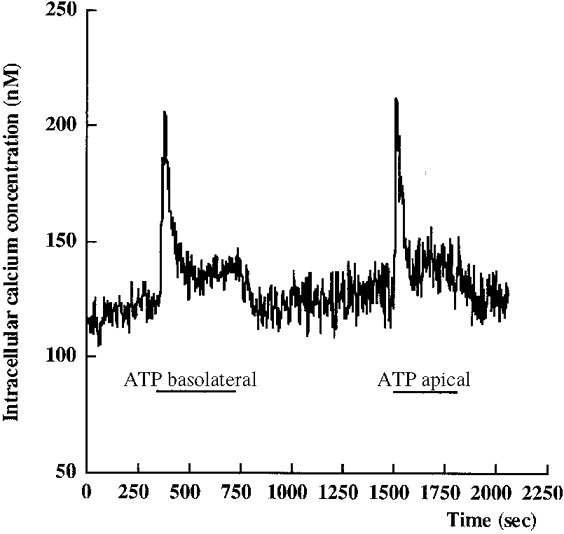
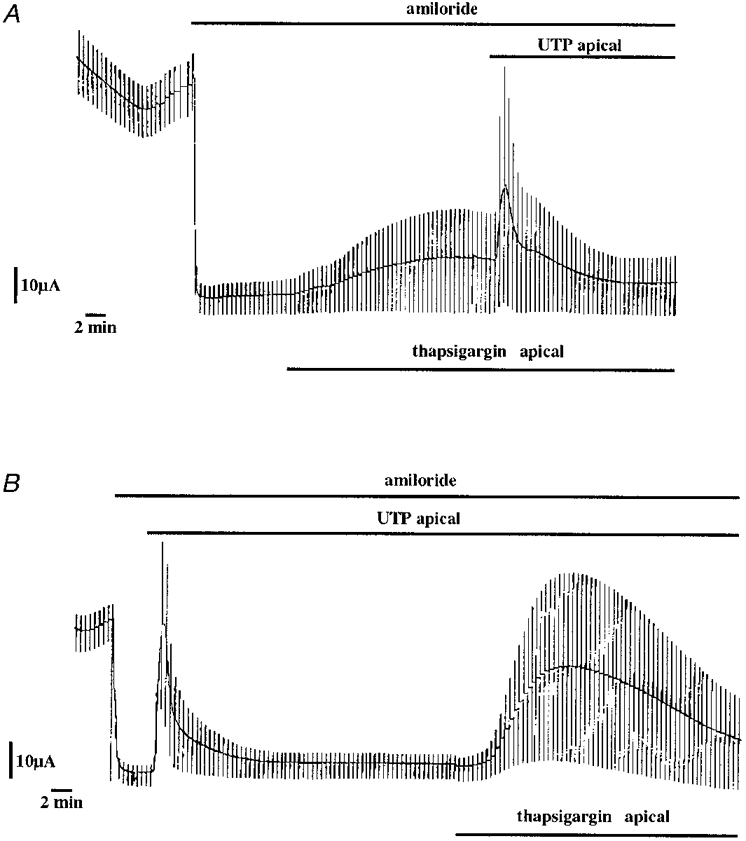
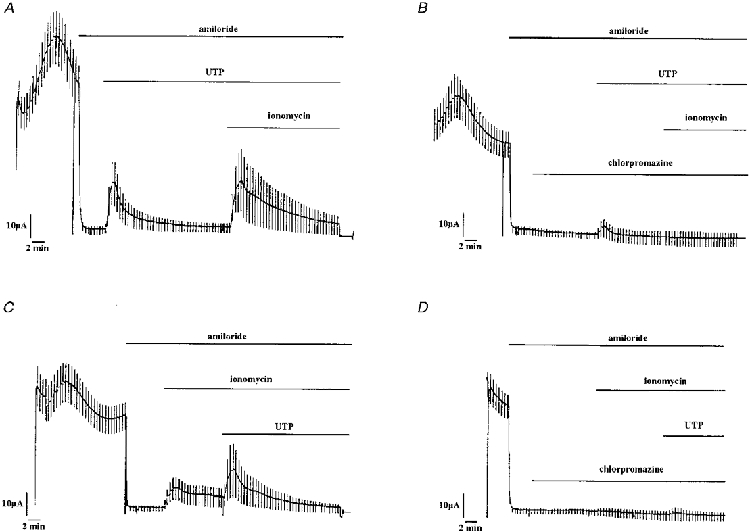
1. The effect of extracellular nucleotides applied on the apical side of polarised A6 cells grown on permeant filters was investigated by measuring the changes in (i) the 36Cl efflux through the apical membranes, (ii) the intracellular chloride concentrations (aCli, measured with N-(6-methoxyquinolyl) acetoethyl ester, MQAE), (iii) ICl, the short-circuit current in the absence of Na+ transport and (iv) the characteristics of the apical chloride channels using a patch-clamp approach. 2. ATP or UTP (0.1-500 microM) transiently stimulated ICl. The sequence of purinergic agonist potencies was UTP = ATP > ADP >> the P2X-selective agonist beta,gamma-methylene ATP = the P2Y-selective agonist 2-methylthioATP. Suramin (100 microM) as the P2Y antagonist Reactive Blue 2 (10 microM) had no effect on the UTP (or ATP)-stimulated current. These findings are consistent with the presence of P2Y2-like receptors located on the apical membranes of A6 cells. Apical application of adenosine also transiently increased ICl. This effect was blocked by theophylline while the UTP-stimulated ICl was not. The existence of a second receptor, of the P1 type is proposed. 3. ATP (or UTP)-stimulated ICl was blocked by apical application of 200 microM N-phenylanthranilic acid (DPC) or 100 microM niflumic acid while 100 microM glibenclamide was ineffective. 4. Ionomycin and thapsigargin both transiently stimulated ICl; the nucleotide stimulation of ICl was not suppressed by pre-treatment with these agents. Chlorpromazin (50 microM), a Ca2+-calmodulin inhibitor strongly inhibited the stimulation of ICl induced either by apical UTP or by ionomycin application. BAPTA-AM pre-treatment of A6 cells blocked the UTP-stimulated ICl. Niflumic acid also blocked the ionomycin stimulated ICl. 5. A fourfold increase in 36Cl effluxes through the apical membranes was observed after ATP or UTP application. These increases of the apical chloride permeability could also be observed when following aCli changes. Apical application of DPC (1 mM) or 5-nitro-2(3-phenylpropylamino)benzoic acid (NPPB; 500 microM) produced an incomplete inhibition of 36Cl effluxes through the apical membranes in ATP-stimulated and in untreated monolayers. 6. In single channel patch-clamp experiments, an apical chloride channel with a unitary single channel conductance of 7.3 +/- 0.6 pS (n = 12) was usually observed. ATP application induced the activation of one or more of these channels within a few minutes. 7. These results indicate that multiple purinergic receptor subtypes are present in the apical membranes of A6 cells and that nucleotides can act as modulators of Cl- secretion in renal cells.
Figures
References
-
- Barnard EA, Simon J, Webb TE. Nucleotide receptors in the nervous system. An abundant component using diverse transduction mechanisms. Review of Molecular Neurobiology. 1997;15:103–129. - PubMed
-
- Begenisich T, Melvin JE. Regulation of chloride channels in epithelia. Journal of Membrane Biology. 1998;163:77–85. - PubMed
-
- Bhagwat SS, Williams M. Purine and pyrimidine receptors: emerging superfamilies of G-protein-coupled and ligand-gated-ion channel receptors. European Journal of Medical Chemistry. 1997;32:183–193.
-
- Brochiero E, Banderali U, Lindenthal S, Raschi C, Ehrenfeld J. Basolateral membrane chloride permeability of A6 cells: implication in cell volume regulation. Pflügers Archiv. 1995a;431:32–46. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous