

Formation of compact myelin is required for maturation of the axonal cytoskeleton
- PMID: 10460234
- PMCID: PMC6782532
- DOI: 10.1523/JNEUROSCI.19-17-07278.1999
Formation of compact myelin is required for maturation of the axonal cytoskeleton
Abstract
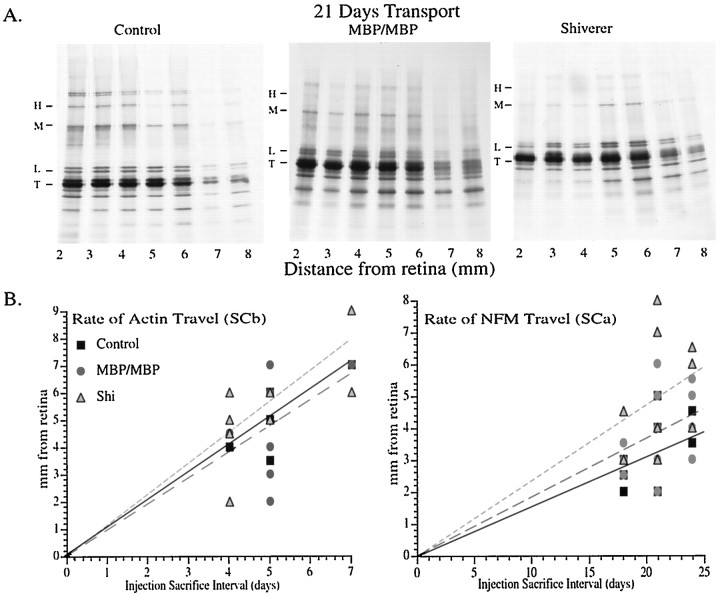
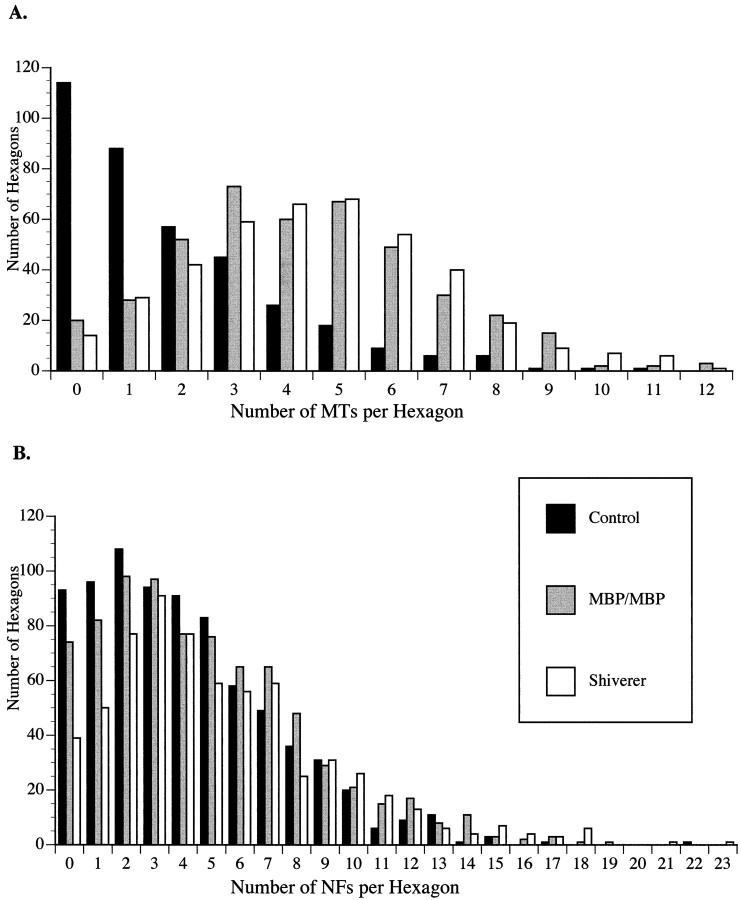
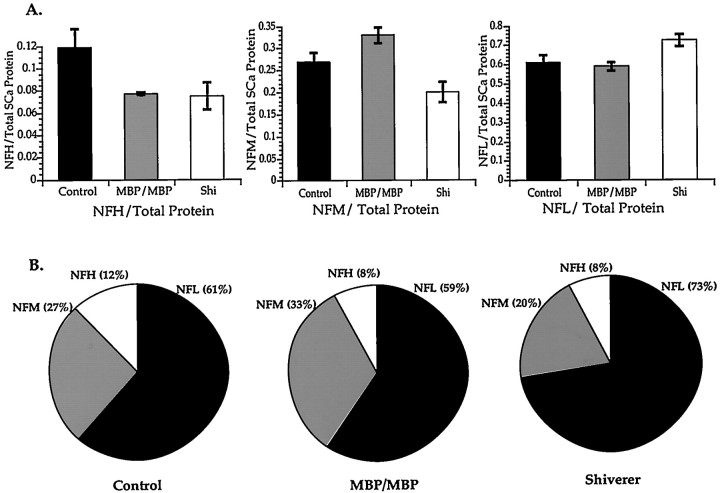
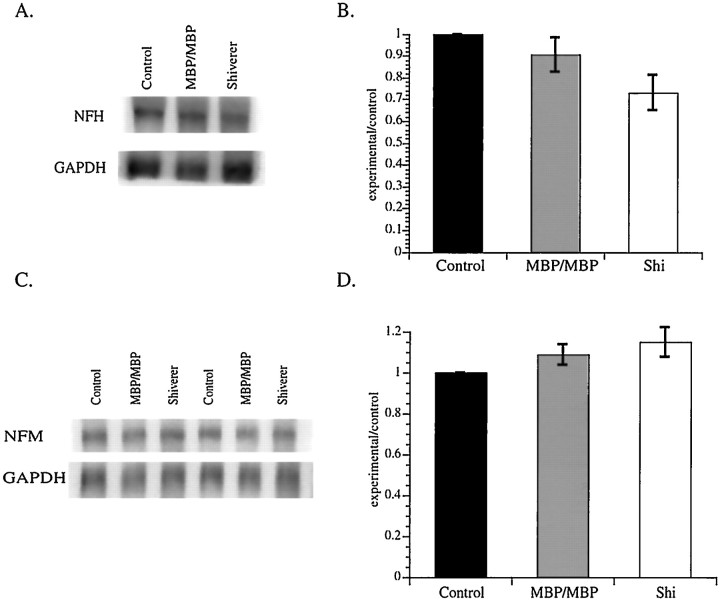
Although traditional roles ascribed to myelinating glial cells are structural and supportive, the importance of compact myelin for proper functioning of the nervous system can be inferred from mutations in myelin proteins and neuropathologies associated with loss of myelin. Myelinating Schwann cells are known to affect local properties of peripheral axons (de Waegh et al., 1992), but little is known about effects of oligodendrocytes on CNS axons. The shiverer mutant mouse has a deletion in the myelin basic protein gene that eliminates compact myelin in the CNS. In shiverer mice, both local axonal features like phosphorylation of cytoskeletal proteins and neuronal perikaryon functions like cytoskeletal gene expression are altered. This leads to changes in the organization and composition of the axonal cytoskeleton in shiverer unmyelinated axons relative to age-matched wild-type myelinated fibers, although connectivity and patterns of neuronal activity are comparable. Remarkably, transgenic shiverer mice with thin myelin sheaths display an intermediate phenotype indicating that CNS neurons are sensitive to myelin sheath thickness. These results indicate that formation of a normal compact myelin sheath is required for normal maturation of the neuronal cytoskeleton in large CNS neurons.
Figures



References
-
- Aguayo A, Attiwell M, Trecarten J, Perkins S, Bray G. Abnormal myelination in transplanted Trembler mouse Schwann cells. Nature. 1977;265:73–74. - PubMed
-
- Brady S. Axonal transport methods and applications. In: Boulton A, Baker G, editors. Neuromethods, general neurochemical techniques. Humana; Clifton, NJ: 1985. pp. 419–476.
-
- Brady ST. Axonal dynamics and regeneration. In: Gorio A, editor. Neuroregeneration. Raven; New York: 1993. pp. 7–36.
-
- Campagnoni AT, Macklin WB. Cellular and molecular aspects of myelin protein gene expression. Mol Neurobiol. 1988;2:41–89. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases