

Specificity and sensitivity of a human olfactory receptor functionally expressed in human embryonic kidney 293 cells and Xenopus Laevis oocytes
- PMID: 10460249
- PMCID: PMC6782526
- DOI: 10.1523/JNEUROSCI.19-17-07426.1999
Specificity and sensitivity of a human olfactory receptor functionally expressed in human embryonic kidney 293 cells and Xenopus Laevis oocytes
Abstract
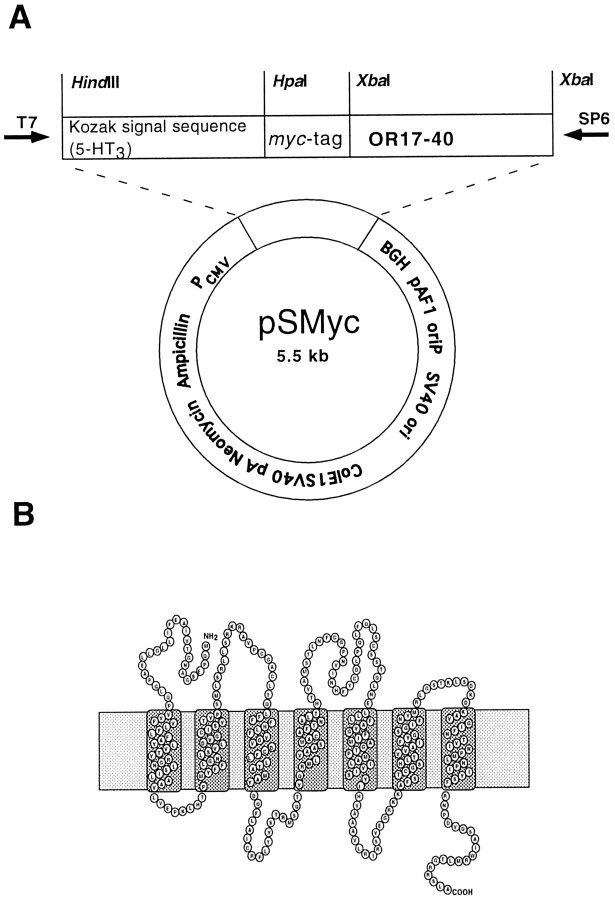
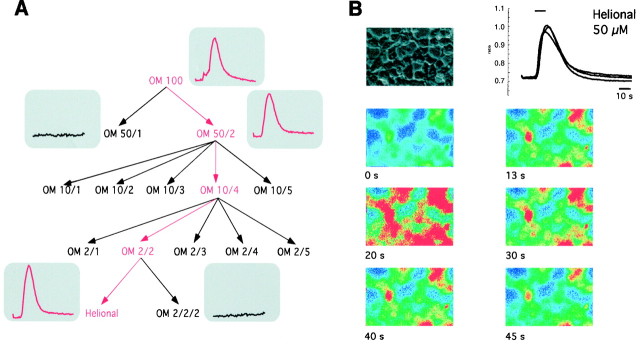
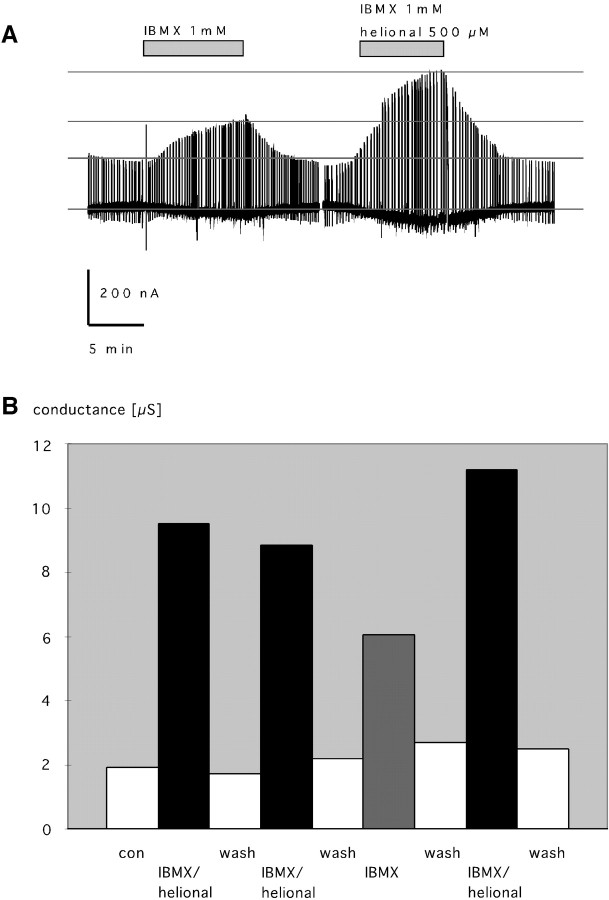
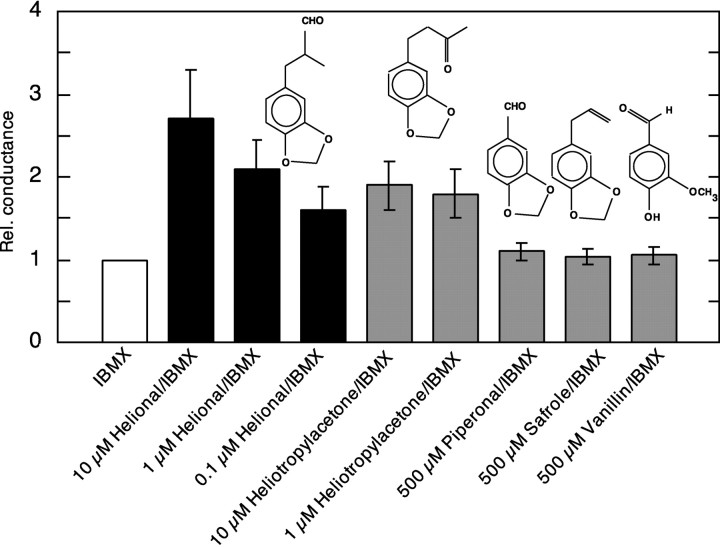
Here, we provide the first evidence for functional expression of a human olfactory receptor protein (OR17-40) and show that recombinant olfactory receptors can be functionally expressed in heterologous systems. A mixture of 100 different odorants (Henkel 100) elicited a transient increase in intracellular [Ca(2+)] in human embryonic kidney 293 (HEK293) cells stably or transiently transfected with the plasmid pOR17-40. By subdividing the odorant mixture into progressively smaller groups, we identified a single component that represented the only effective substance: helional. Only the structurally closely related molecule heliotroplyacetone also activated the receptor. Other compounds, including piperonal, safrole, and vanillin, were completely ineffective. Mock-transfected cells and cells transfected with other receptors showed no change in intracellular [Ca(2+)] in response to odor stimulation. We were also able to functionally express OR17-40 in Xenopus laevis oocytes. Coexpression of a "reporter" channel allowed measurement of the response of oocytes injected with the cRNA of the human receptor to the odor mixture Henkel 100. The effective substances were the same (helional, heliotropylacetone) as those identified by functionally expressing the receptor in HEK293 cells and were active at the same, lower micromolar concentration. These findings open the possibility of now characterizing the sensitivity and specificity of many, if not all, of the hundreds of different human olfactory receptors.
Figures




References
-
- Afshar M, Hubbard RE, Demaille J. Towards structural models of molecular recognition in olfactory receptors. Biochimie. 1998;80:129–135. - PubMed
-
- Axel R. The molecular logic of smell. Sci Am. 1995;273:154–159. - PubMed
-
- Beets J. Olfactory response and molecular structure. Handbook of sensory physiology 1971. 4:257–321.
-
- Ben-Arie N, Lancet D, Taylor C, Khen M, Walker N, Carrozzo R, Patel K, Sheer D, Lehrach H, North MA. Olfactory receptor gene cluster on human chromosome 17: possible duplication of an ancestral receptor repertoire. Hum Mol Genet. 1994;3:229–235. - PubMed
-
- Buck L, Axel R. A novel multigene family may encode odorant receptors: a molecular basis for odor recognition. Cell. 1991;65:175–187. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous