

Effects of attention on the processing of motion in macaque middle temporal and medial superior temporal visual cortical areas
- PMID: 10460265
- PMCID: PMC6782504
- DOI: 10.1523/JNEUROSCI.19-17-07591.1999
Effects of attention on the processing of motion in macaque middle temporal and medial superior temporal visual cortical areas
Abstract
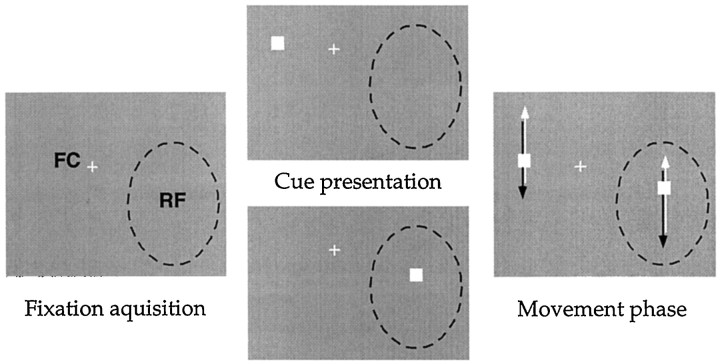
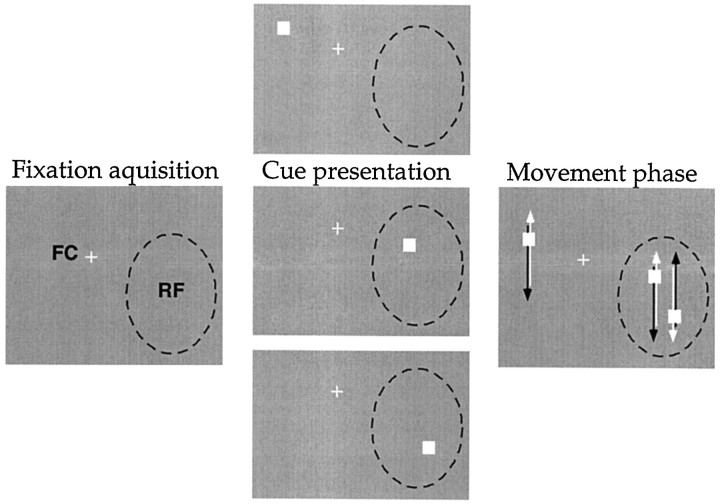
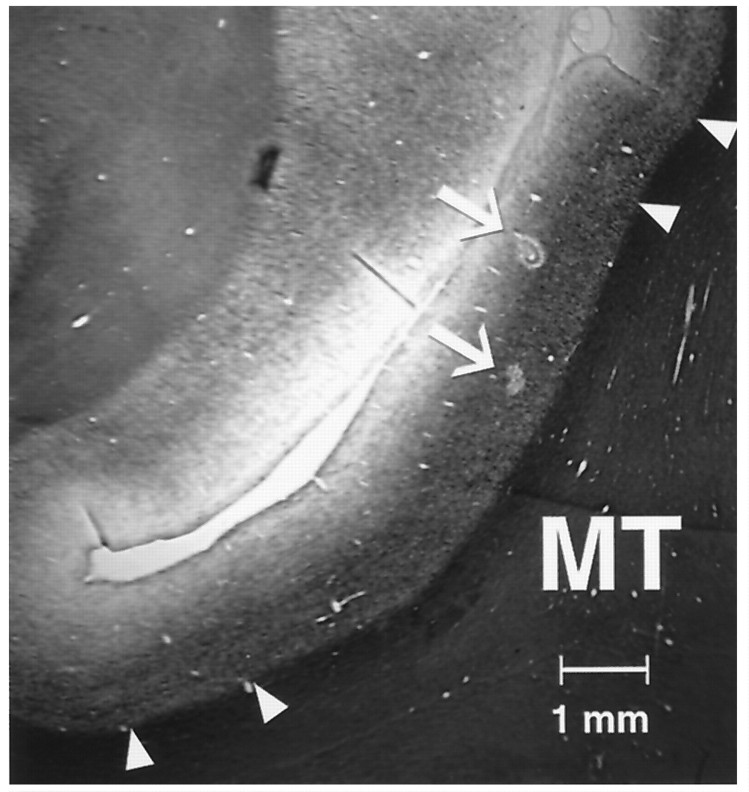
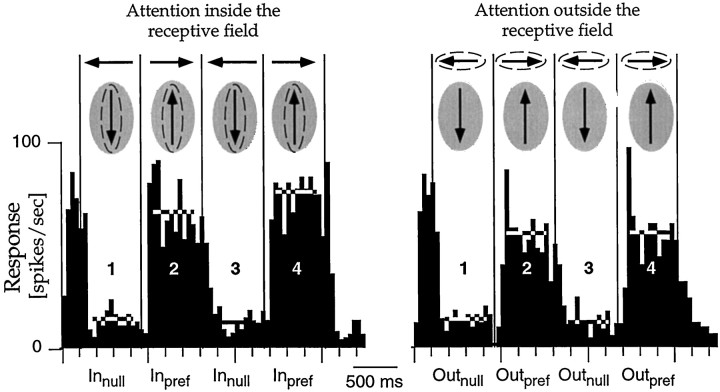
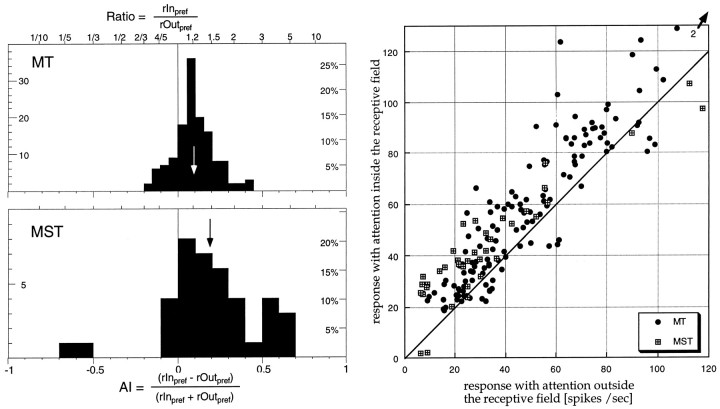
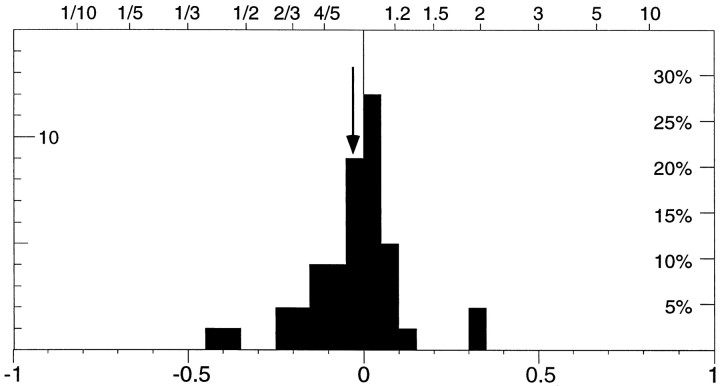
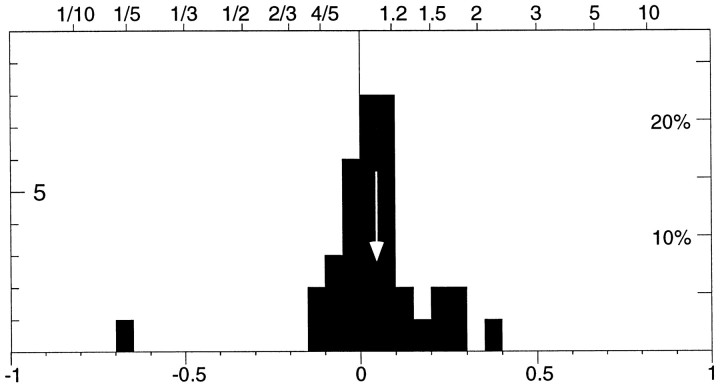
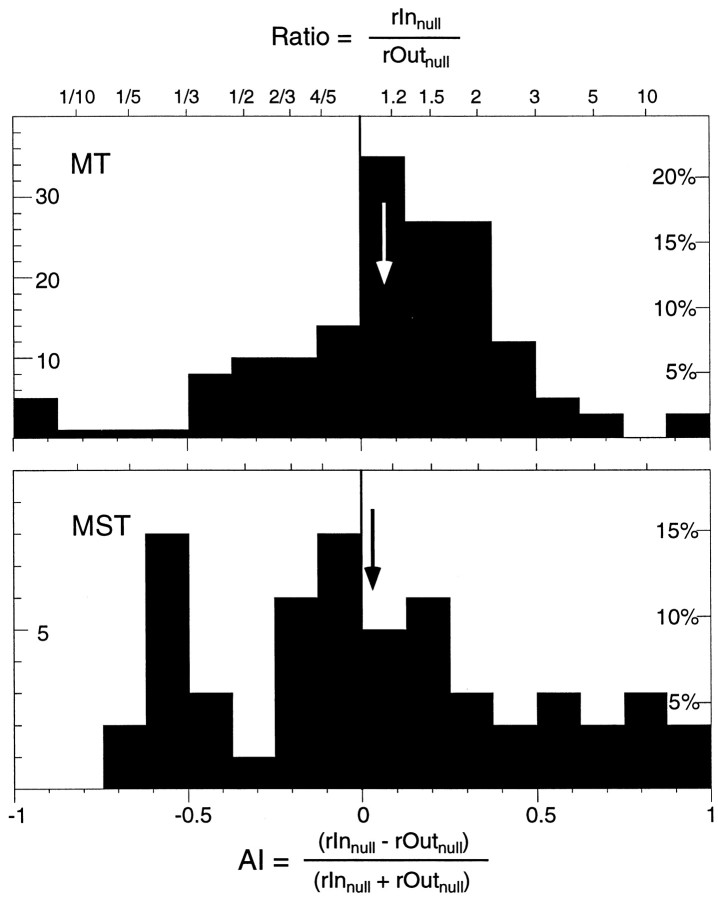
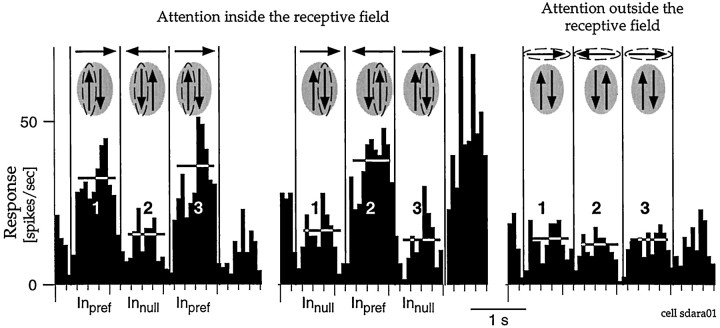
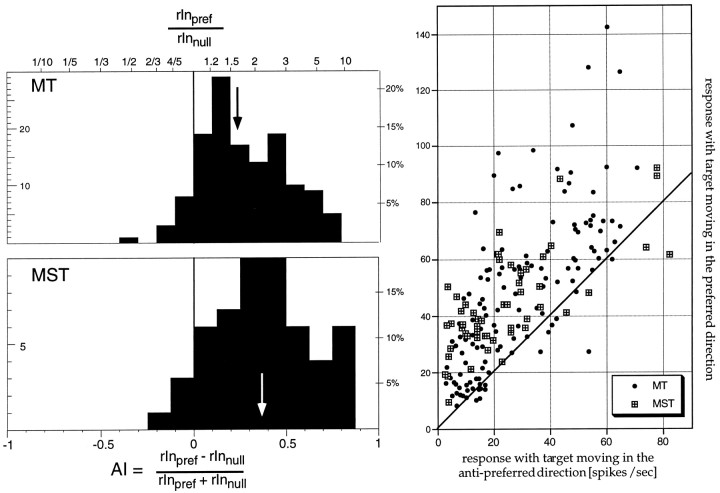
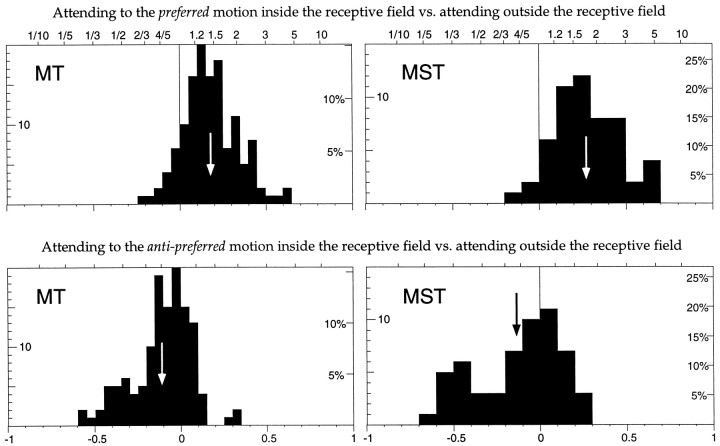
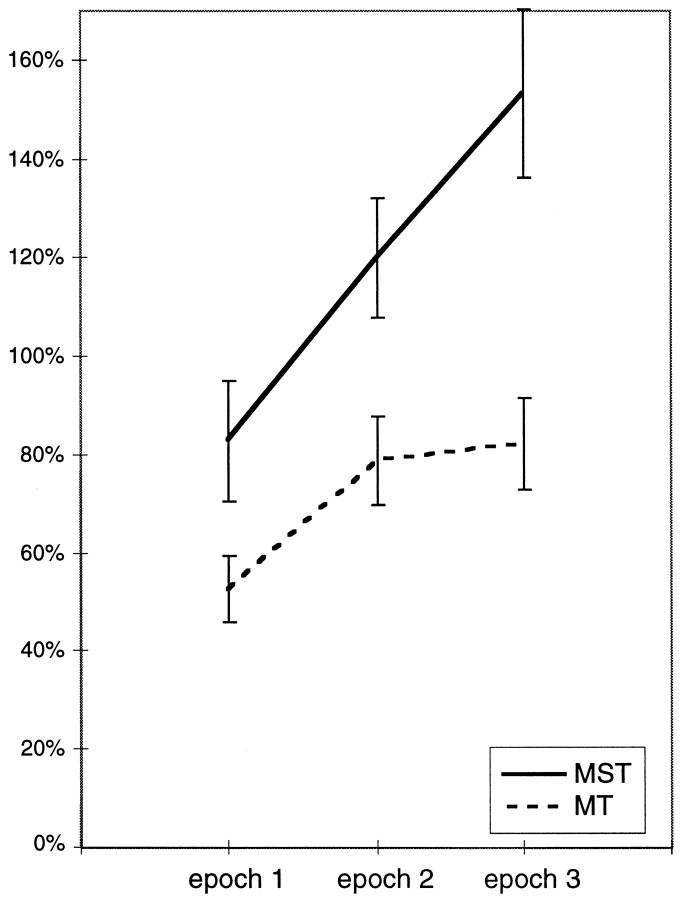
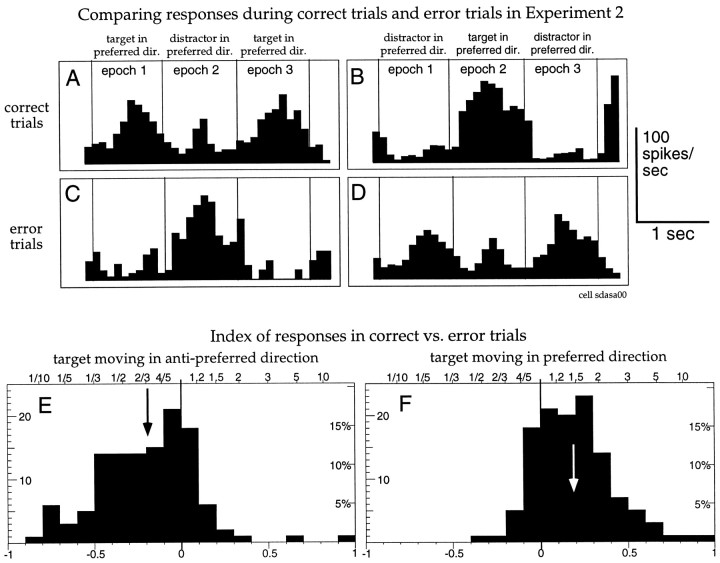
The visual system is continually inundated with information received by the eyes. Only a fraction of this information appears to reach visual awareness. This process of selection is one of the functions ascribed to visual attention. Although many studies have investigated the role of attention in shaping neuronal representations in cortical areas, few have focused on attentional modulation of neuronal signals related to visual motion. We recorded from 89 direction-selective neurons in middle temporal (MT) and medial superior temporal (MST) visual cortical areas of two macaque monkeys using identical sensory stimulation under various attentional conditions. Neural responses in both areas were greatly influenced by attention. When attention was directed to a stimulus inside the receptive field of a neuron, responses in MT and MST were enhanced an average of 20 and 40% compared with a condition in which attention was directed outside the receptive field. Even stronger average enhancements (70% in MT and 100% in MST) were observed when attention was switched from a stimulus moving in the nonpreferred direction inside the receptive field to another stimulus in the receptive field that was moving in the preferred direction. These findings show that attention modulates motion processing from stages early in the dorsal visual pathway by selectively enhancing the representation of attended stimuli and simultaneously reducing the influence of unattended stimuli.
Figures
Similar articles
-
Attentional modulation of visual motion processing in cortical areas MT and MST.Nature. 1996 Aug 8;382(6591):539-41. doi: 10.1038/382539a0. Nature. 1996. PMID: 8700227
-
Responses of MT and MST neurons to one and two moving objects in the receptive field.J Neurophysiol. 1997 Dec;78(6):2904-15. doi: 10.1152/jn.1997.78.6.2904. J Neurophysiol. 1997. PMID: 9405511
-
Feature-based attention increases the selectivity of population responses in primate visual cortex.Curr Biol. 2004 May 4;14(9):744-51. doi: 10.1016/j.cub.2004.04.028. Curr Biol. 2004. PMID: 15120065
-
Primate extrastriate cortical area MST: a gateway between sensation and cognition.J Neurophysiol. 2021 May 1;125(5):1851-1882. doi: 10.1152/jn.00384.2020. Epub 2021 Mar 3. J Neurophysiol. 2021. PMID: 33656951 Review.
-
A neural-based code for computing image velocity from small sets of middle temporal (MT/V5) neuron inputs.J Vis. 2012 Aug 1;12(8):1. doi: 10.1167/12.8.1. J Vis. 2012. PMID: 22854102 Review.
Cited by
-
Relating a Spiking Neural Network Model and the Diffusion Model of Decision-Making.Comput Brain Behav. 2022 Sep;5(3):279-301. doi: 10.1007/s42113-022-00143-4. Epub 2022 Jun 13. Comput Brain Behav. 2022. PMID: 36408474 Free PMC article.
-
Attention selectively modifies the representation of individual faces in the human brain.J Neurosci. 2013 Apr 17;33(16):6979-89. doi: 10.1523/JNEUROSCI.4142-12.2013. J Neurosci. 2013. PMID: 23595755 Free PMC article.
-
Stimulus-specific competitive selection in macaque extrastriate visual area V4.Proc Natl Acad Sci U S A. 2007 Mar 6;104(10):4165-9. doi: 10.1073/pnas.0611722104. Epub 2007 Feb 28. Proc Natl Acad Sci U S A. 2007. PMID: 17360494 Free PMC article.
-
Comparison of the spatial limits on direction selectivity in visual areas MT and V1.J Neurophysiol. 2005 Mar;93(3):1235-45. doi: 10.1152/jn.00767.2004. Epub 2004 Oct 13. J Neurophysiol. 2005. PMID: 15483064 Free PMC article.
-
MECHANISMS OF PERCEPTUAL LEARNING.Learn Percept. 2009 Jun 1;1(1):19-36. doi: 10.1556/LP.1.2009.1.3. Learn Percept. 2009. PMID: 20445764 Free PMC article.
References
-
- Andersen RA, Graziano MSA, Snowden R. Translational invariance and attentional modulation of MST cells. Soc Neurosci Abstr. 1990;16:7.
-
- Assad JA, Maunsell JHR. Neuronal correlates of inferred motion in primate posterior parietal cortex. Nature. 1994;373:518–521. - PubMed
-
- Beauchamp MS, Cox RW, DeYoe EA. Graded effects of spatial and featural attention on human area MT and associated motion processing areas. J Neurophysiol. 1997;78:516–520. - PubMed
-
- Britten KH. Spatial interactions within monkey middle temporal (MT) receptive fields. Soc Neurosci Abstr. 1995;21:663.
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources