

The cytoplasmic Purkinje onconeural antigen cdr2 down-regulates c-Myc function: implications for neuronal and tumor cell survival
- PMID: 10465786
- PMCID: PMC316965
- DOI: 10.1101/gad.13.16.2087
The cytoplasmic Purkinje onconeural antigen cdr2 down-regulates c-Myc function: implications for neuronal and tumor cell survival
Abstract
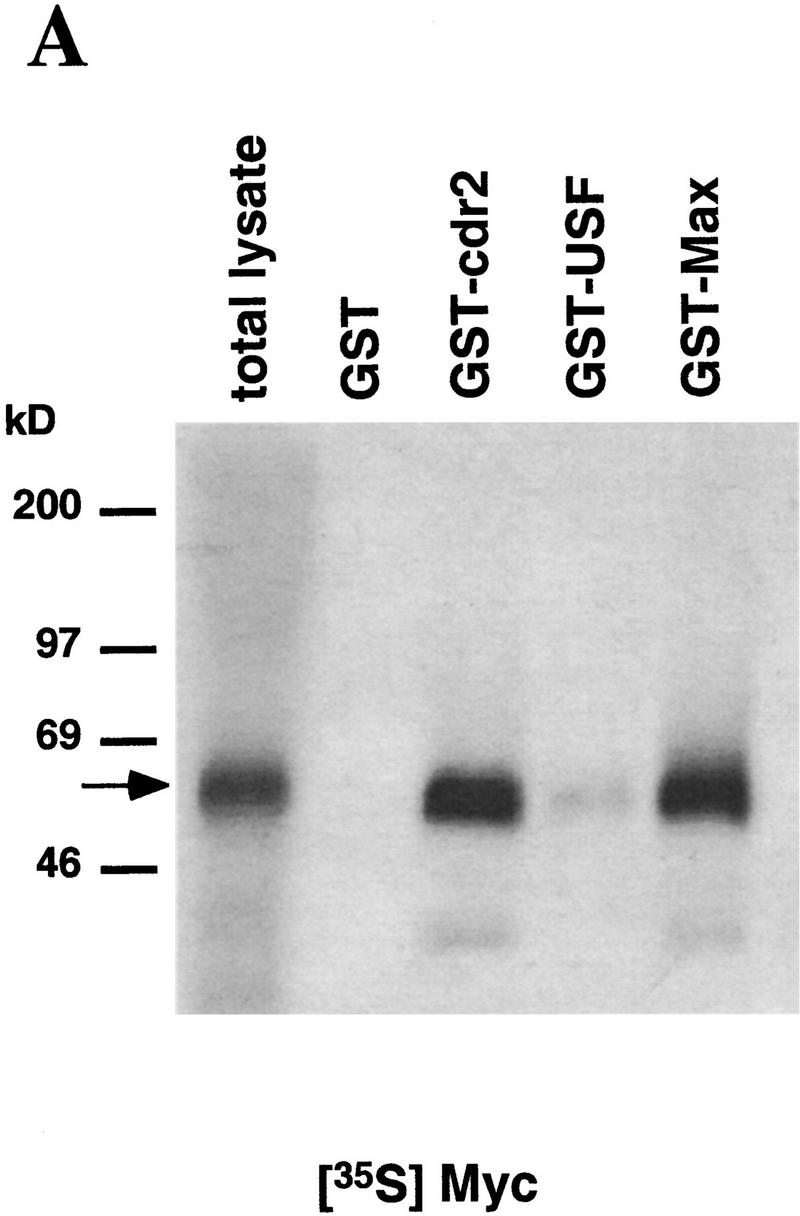
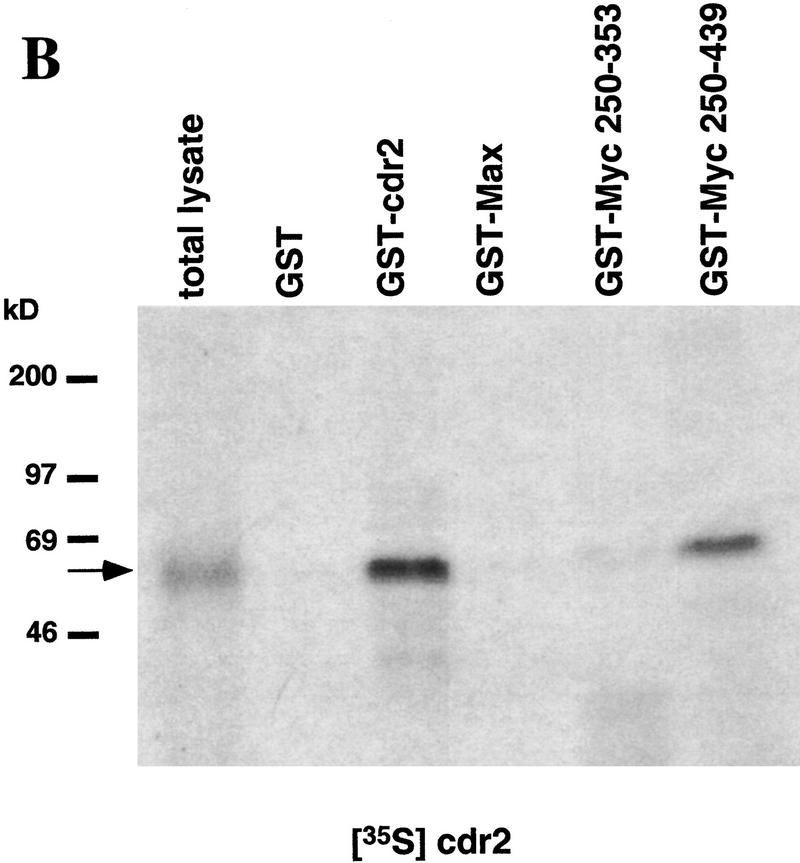
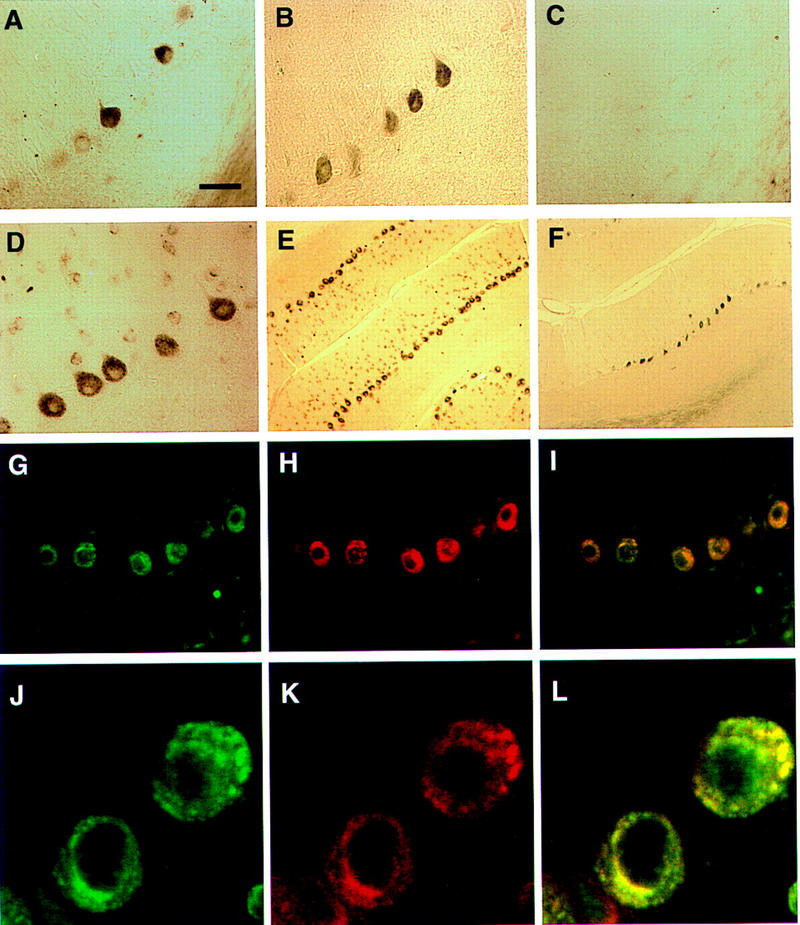
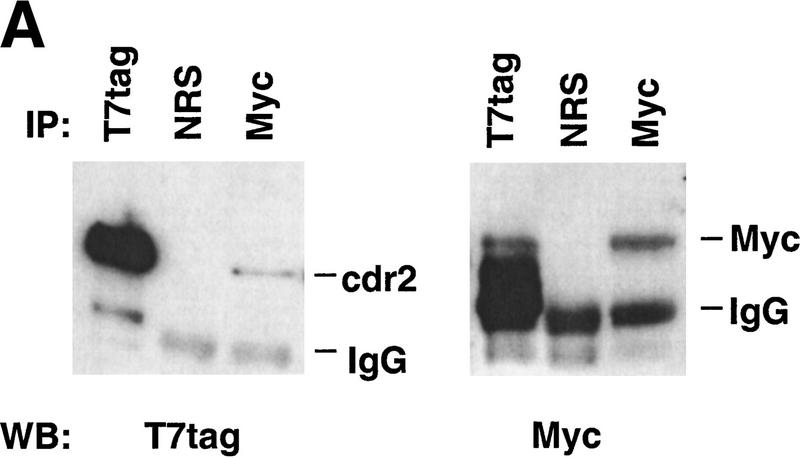
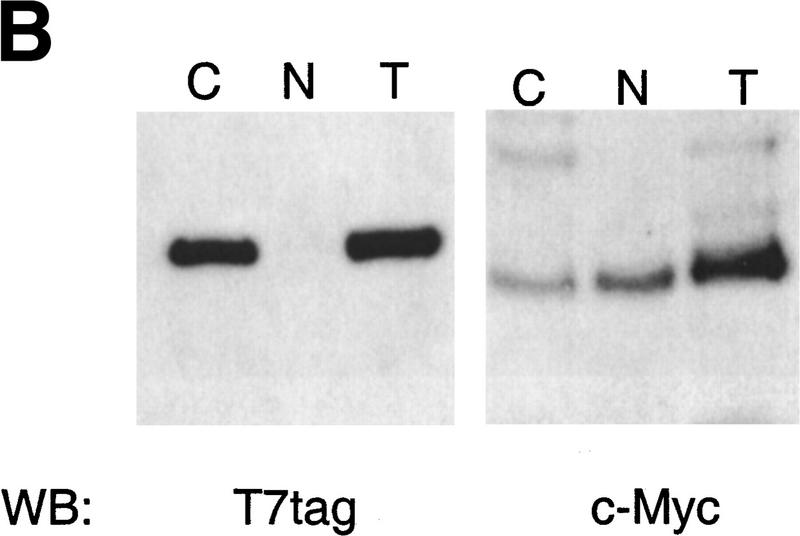
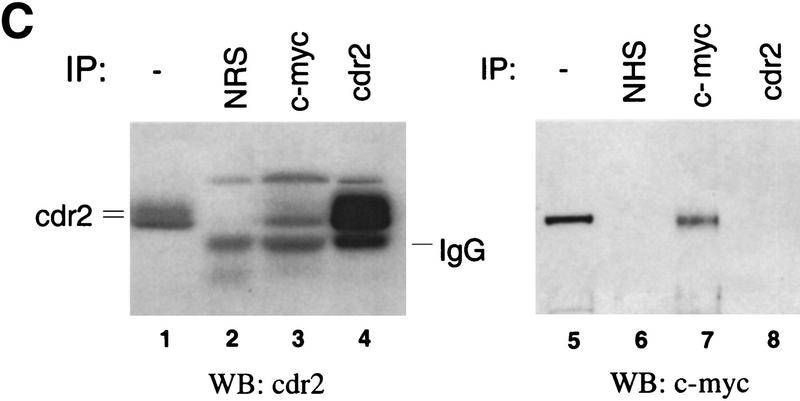
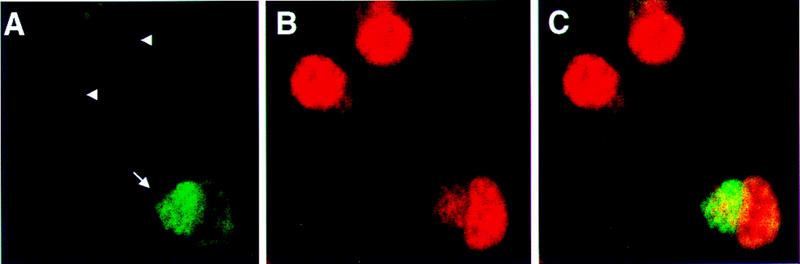
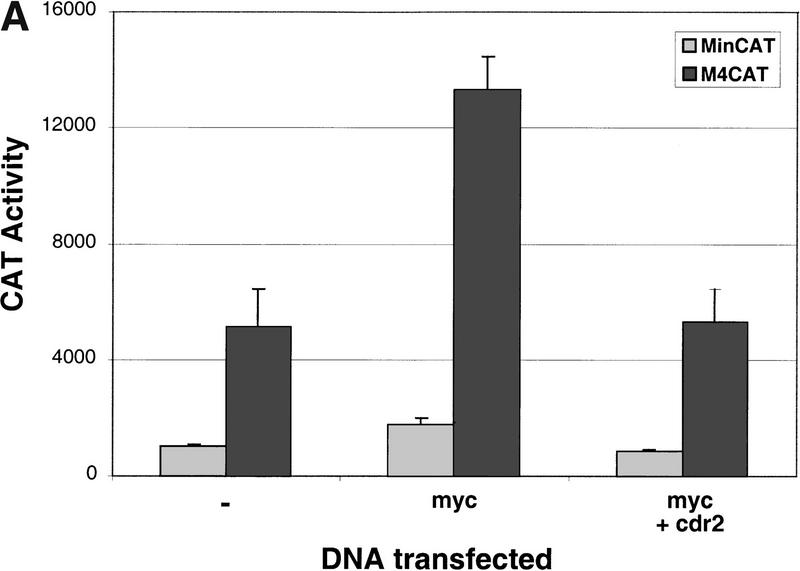
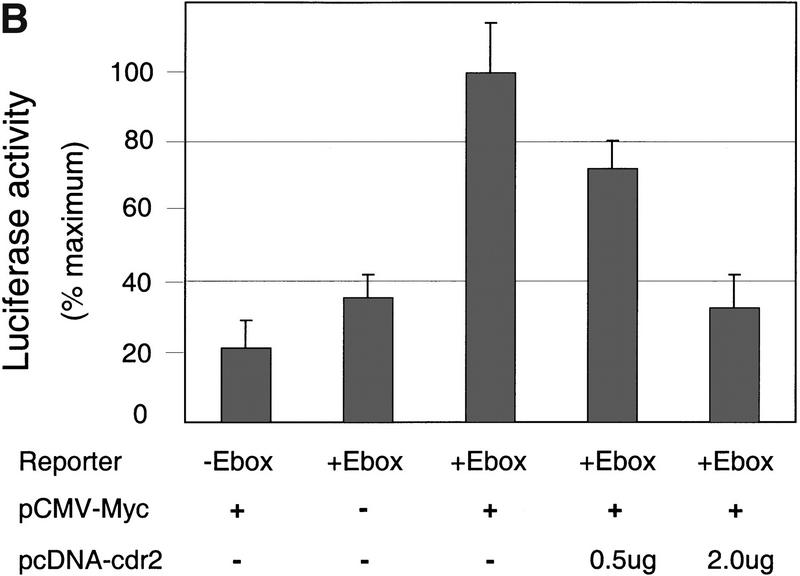
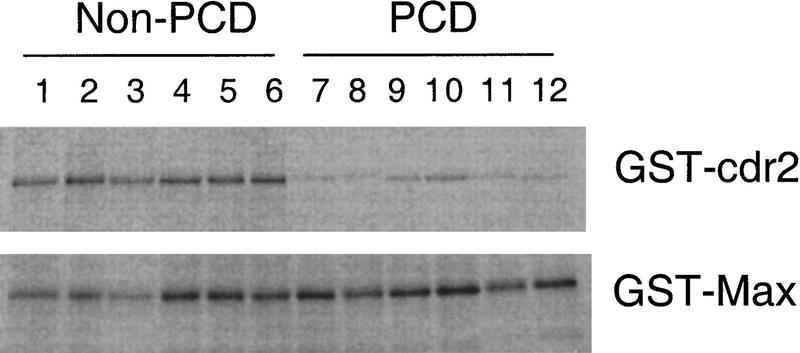
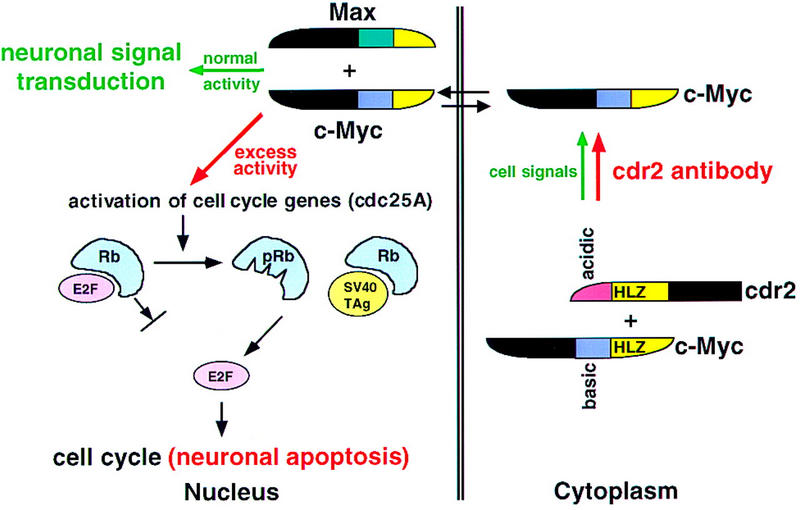
Paraneoplastic cerebellar degeneration (PCD) is a disorder in which breast or ovarian tumors express an onconeural antigen termed cdr2, which normally is expressed in cerebellar Purkinje neurons. This leads to an immune response to cdr2 that is associated with tumor immunity and autoimmune cerebellar degeneration. We have found that cdr2, a cytoplasmic protein harboring a helix-leucine zipper (HLZ) motif, interacts specifically with the HLZ motif of c-Myc. Both proteins colocalize in the cytoplasm of adult cerebellar Purkinje neurons, and coimmunoprecipitate from tumor cell lines and cerebellar extracts. cdr2 down-regulates c-Myc-dependent transcription in cotransfection assays, and redistributes Myc protein in the cytoplasm. Disease antisera from six of six PCD patients specifically blocked the interaction between cdr2 and c-Myc in vitro. These data indicate that cdr2 normally sequesters c-Myc in the neuronal cytoplasm, thereby down-regulating c-Myc activity, and suggest a mechanism whereby inhibition of cdr2 function by autoantibodies in PCD may contribute to Purkinje neuronal death.
Figures
References
-
- Albert ML, Darnell JC, Bender A, Francisco LM, Bhardwaj N, Darnell RB. Tumor-specific killer cells in paraneoplastic cerebellar degeneration. Nature Med. 1998;4:1321–1324. - PubMed
-
- Ayer DE, Kretzner L, Eisenman RN. Mad: A heterodimeric partner for Max that antagonizes Myc transcriptional activity. Cell. 1993;72:211–222. - PubMed
-
- Bai MK, Costopoulos JS, Christoforidou BP, Papadimitriou CS. Immunohistochemical detection of the c-myc oncogene product in normal, hyperplastic and carcinomatous endometrium. Oncology. 1994;51:314–319. - PubMed
-
- Bao J, Zervos AS. Isolation and characterization of Nmi, a novel partner of Myc proteins. Oncogene. 1996;12:2171–2176. - PubMed
-
- Blackwood EM, Luscher B, Eisenman RN. Myc and Max associate in vivo. Genes & Dev. 1992;6:71–80. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases