

Role of calcium permeation in dihydropyridine receptor function. Insights into channel gating and excitation-contraction coupling
- PMID: 10469729
- PMCID: PMC2229453
- DOI: 10.1085/jgp.114.3.393
Role of calcium permeation in dihydropyridine receptor function. Insights into channel gating and excitation-contraction coupling
Abstract
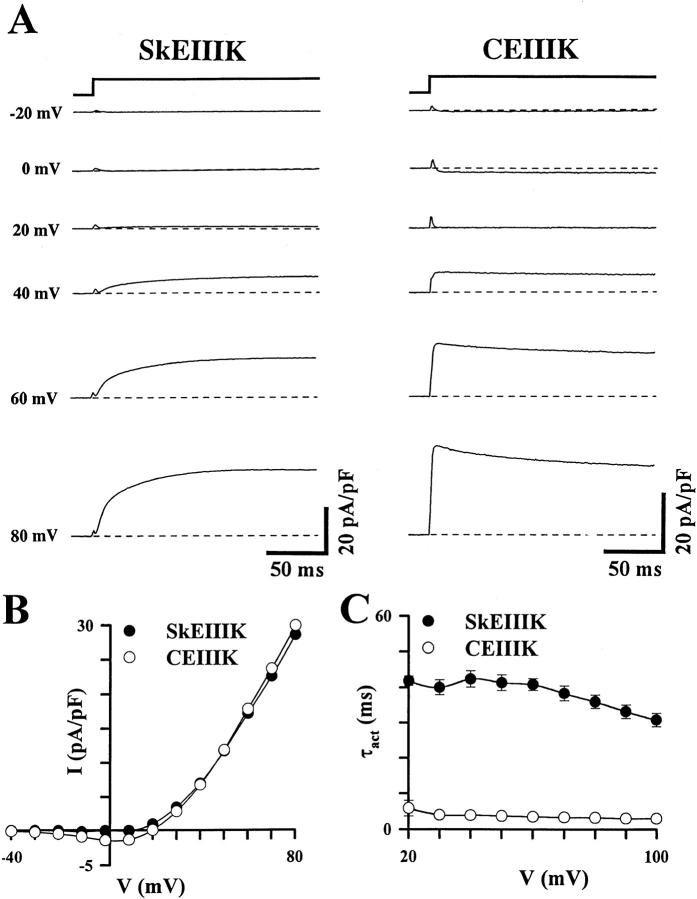
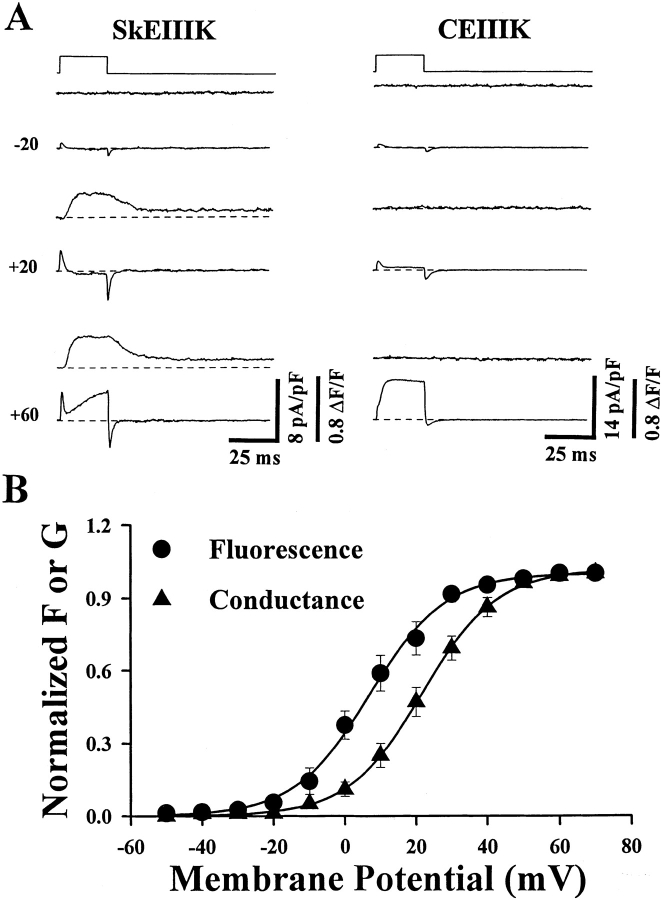
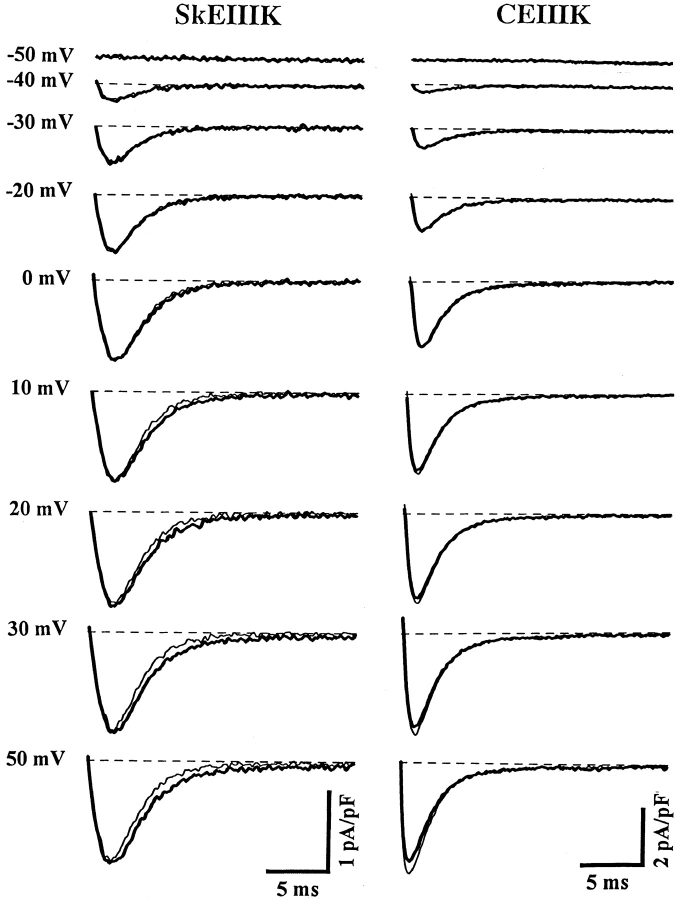
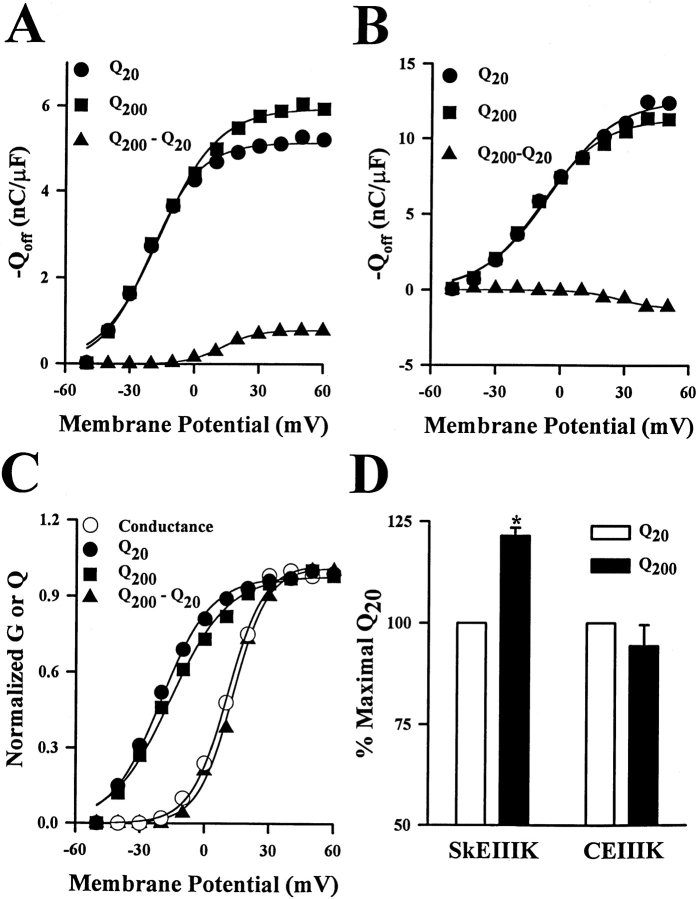
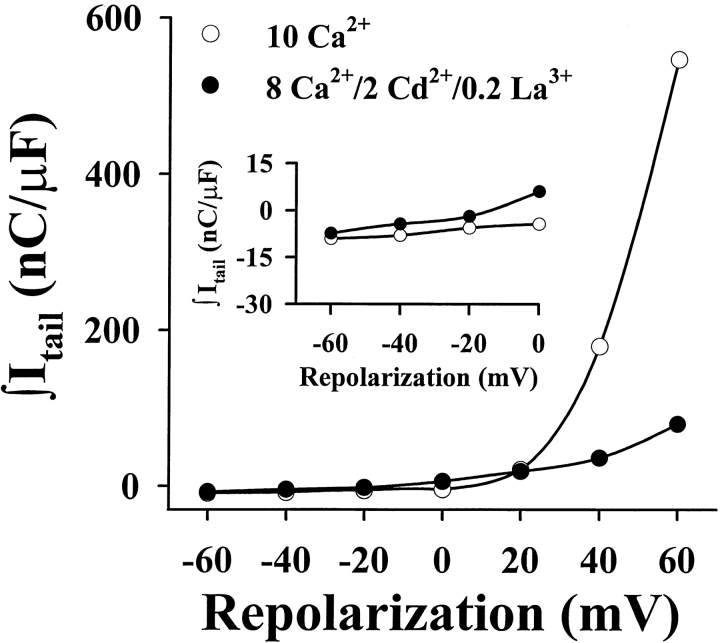
The skeletal and cardiac muscle dihydropyridine receptors (DHPRs) differ with respect to their rates of channel activation and in the means by which they control Ca2+ release from the sarcoplasmic reticulum (Adams, B.A., and K.G. Beam. 1990. FASEB J. 4:2809-2816). We have examined the functional properties of skeletal (SkEIIIK) and cardiac (CEIIIK) DHPRs in which a highly conserved glutamate residue in the pore region of repeat III was mutated to a positively charged lysine residue. Using expression in dysgenic myotubes, we have characterized macroscopic ionic currents, intramembrane gating currents, and intracellular Ca2+ transients attributable to these two mutant DHPRs. CEIIIK supported very small inward Ca2+ currents at a few potentials (from -20 to +20 mV) and large outward cesium currents at potentials greater than +20 mV. SkEIIIK failed to support inward Ca2+ flux at any potential. However, large, slowly activating outward cesium currents were observed at all potentials greater than + 20 mV. The difference in skeletal and cardiac Ca2+ channel activation kinetics was conserved for outward currents through CEIIIK and SkEIIIK, even at very depolarized potentials (at +100 mV; SkEIIIK: tau(act) = 30.7 +/- 1.9 ms, n = 11; CEIIIK: tau(act) = 2.9 +/- 0.5 ms, n = 7). Expression of SkEIIIK in dysgenic myotubes restored both evoked contractions and depolarization-dependent intracellular Ca(2+) transients with parameters of voltage dependence (V(0.5) = 6.5 +/- 3.2 mV and k = 9.3 +/- 0.7 mV, n = 5) similar to those for the wild-type DHPR (Garcia, J., T. Tanabe, and K.G. Beam. 1994. J. Gen. Physiol. 103:125-147). However, CEIIIK-expressing myotubes never contracted and failed to exhibit depolarization-dependent intracellular Ca2+ transients at any potential. Thus, high Ca2+ permeation is required for cardiac-type excitation-contraction coupling reconstituted in dysgenic myotubes, but not skeletal-type. The strong rectification of the EIIIK channels made it possible to obtain measurements of gating currents upon repolarization to -50 mV (Qoff) following either brief (20 ms) or long (200 ms) depolarizing pulses to various test potentials. For SkEIIIK, and not CEIIK, Qoff was significantly (P < 0.001) larger after longer depolarizations to +60 mV (121.4 +/- 2.0%, n = 6). The increase in Qoff for long depolarizations exhibited a voltage dependence similar to that of channel activation. Thus, the increase in Q(off) may reflect a voltage sensor movement required for activation of L-type Ca2+ current and suggests that most DHPRs in skeletal muscle undergo this voltage-dependent transition.
Figures
References
-
- Adams B.A., Tanabe T., Mikami A., Numa S., Beam K.G. Intramembrane charge movement restored in dysgenic skeletal muscle by injection of dihydropyridine receptor cDNAs. Nature. 1990;346:569–572. - PubMed
-
- Armstrong C.M., Bezanilla F.M., Horowicz P. Twitches in the presence of ethylene glycol bis(-aminoethyl ether)-N,N′-tetraacetic acid. Biochim. Biophys. Acta. 1972;267:605–608. - PubMed
-
- Beam K.G., Franzini-Armstrong C. Functional and structural approaches to the study of excitation–contraction coupling. Methods Cell Biol. 1997;52:283–306. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
