

Role of Ngamma-acetyldiaminobutyrate as an enzyme stabilizer and an intermediate in the biosynthesis of hydroxyectoine
- PMID: 10473374
- PMCID: PMC99699
- DOI: 10.1128/AEM.65.9.3774-3779.1999
Role of Ngamma-acetyldiaminobutyrate as an enzyme stabilizer and an intermediate in the biosynthesis of hydroxyectoine
Abstract
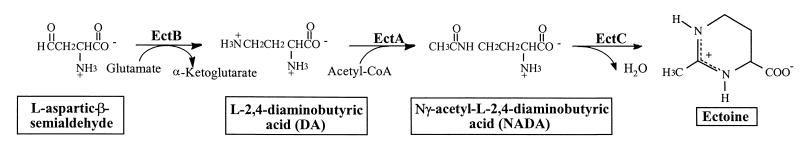
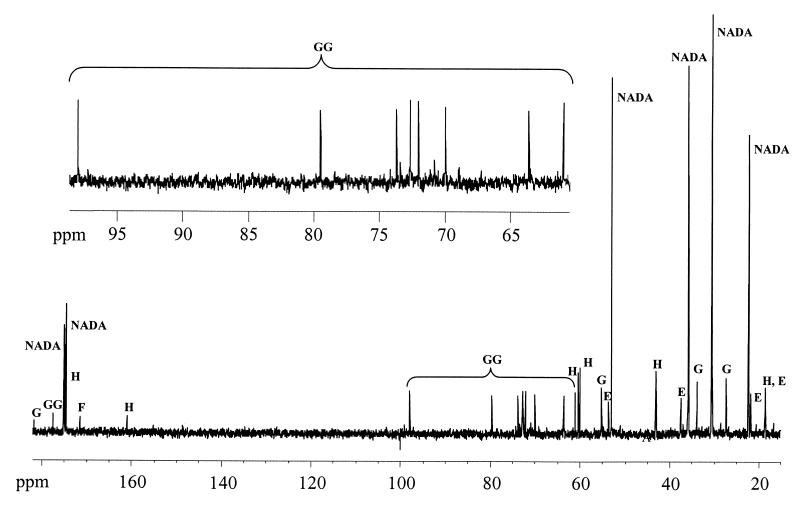
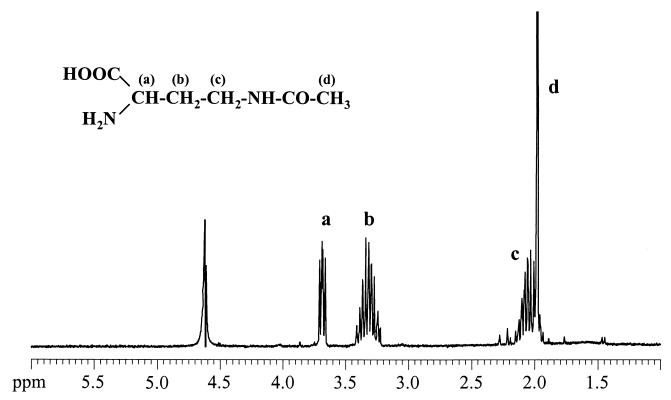
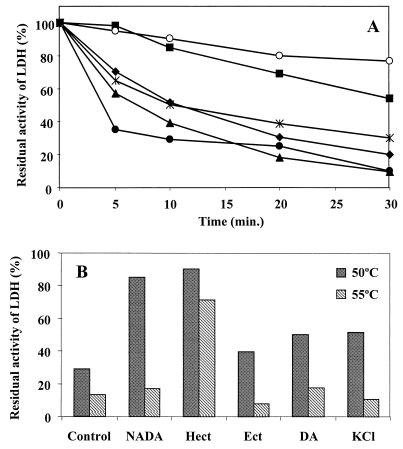
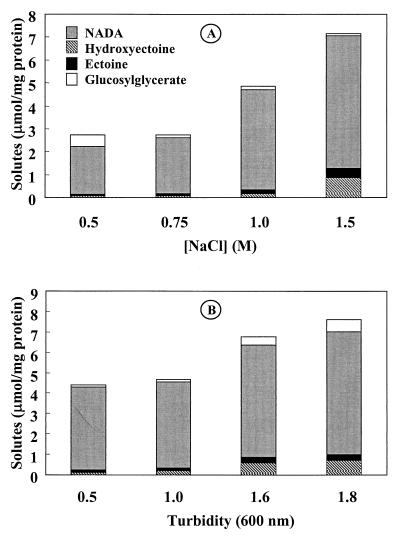
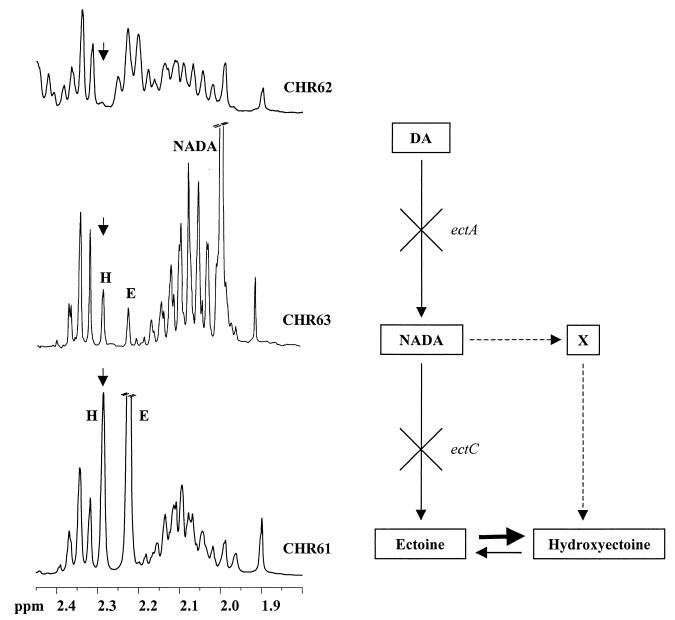
Strain CHR63 is a salt-sensitive mutant of the moderately halophilic wild-type strain Halomonas elongata DSM 3043 that is affected in the ectoine synthase gene (ectC). This strain accumulates large amounts of Ngamma-acetyldiaminobutyrate (NADA), the precursor of ectoine (D. Cánovas, C. Vargas, F. Iglesias-Guerra, L. N. Csonka, D. Rhodes, A. Ventosa, and J. J. Nieto, J. Biol. Chem. 272:25794-25801, 1997). Hydroxyectoine, ectoine, and glucosylglycerate were also identified by nuclear magnetic resonance (NMR) as cytoplasmic organic solutes in this mutant. Accumulation of NADA, hydroxyectoine, and ectoine was osmoregulated, whereas the levels of glucosylglycerate decreased at higher salinities. The effect of the growth stage on the accumulation of solutes was also investigated. NADA was purified from strain CHR63 and was shown to protect the thermolabile enzyme rabbit muscle lactate dehydrogenase against thermal inactivation. The stabilizing effect of NADA was greater than the stabilizing effect of ectoine or potassium diaminobutyrate. A (1)H NMR analysis of the solutes accumulated by the wild-type strain and mutants CHR62 (ectA::Tn1732) and CHR63 (ectC::Tn1732) indicated that H. elongata can synthesize hydroxyectoine by two different pathways-directly from ectoine or via an alternative pathway that converts NADA into hydroxyectoine without the involvement of ectoine.
Figures
References
-
- Bradford M M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal Biochem. 1976;72:248–254. - PubMed
-
- Cánovas D, Vargas C, Calderón M I, Ventosa A, Nieto J J. Characterization of the genes for the biosynthesis of the compatible solute ectoine in the moderately halophilic bacterium Halomonas elongata DSM 3043. Syst Appl Microbiol. 1998;21:487–497. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
