

Stimulation of beta1-integrin function by epidermal growth factor and heregulin-beta has distinct requirements for erbB2 but a similar dependence on phosphoinositide 3-OH kinase
- PMID: 10473632
- PMCID: PMC25524
- DOI: 10.1091/mbc.10.9.2861
Stimulation of beta1-integrin function by epidermal growth factor and heregulin-beta has distinct requirements for erbB2 but a similar dependence on phosphoinositide 3-OH kinase
Abstract
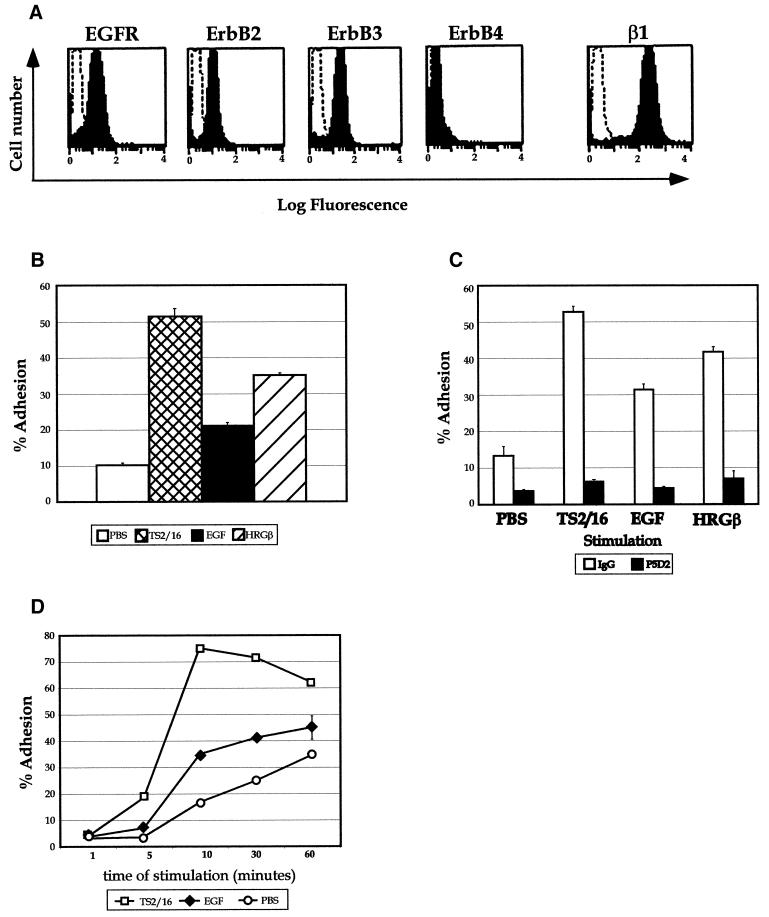
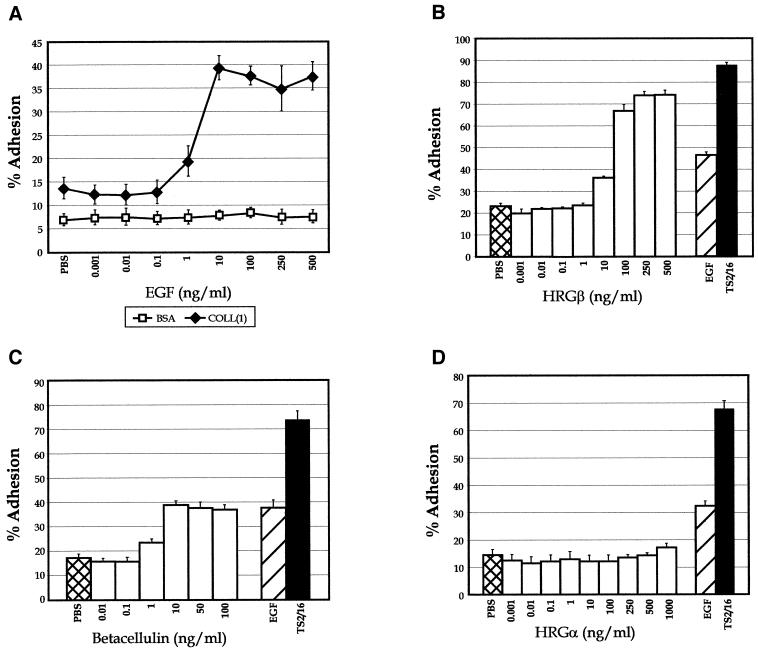
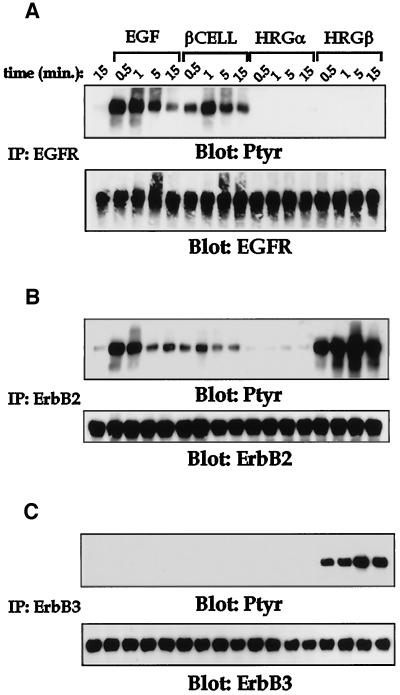
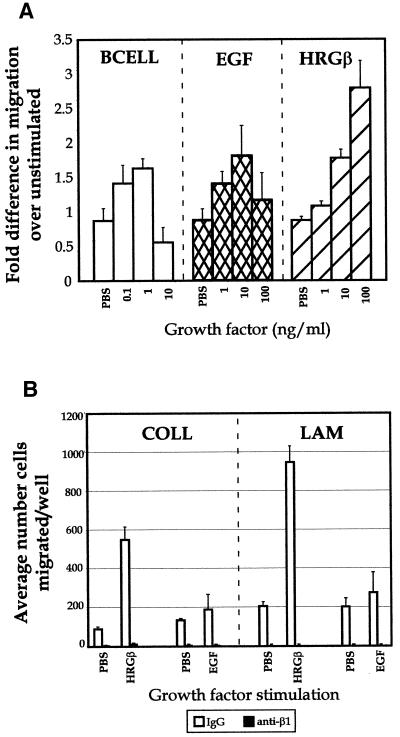
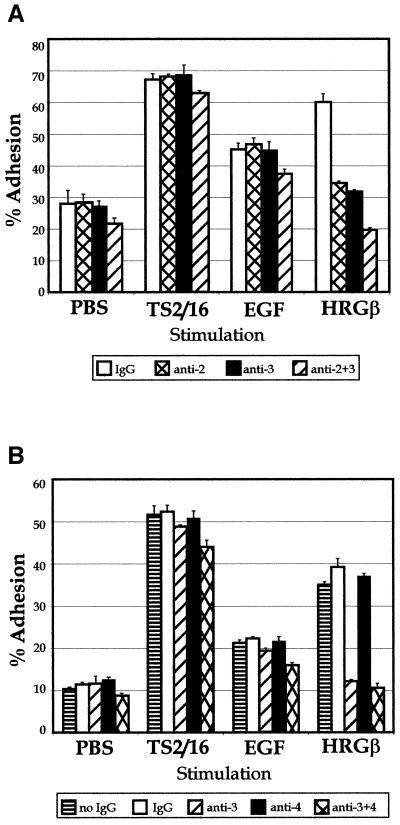
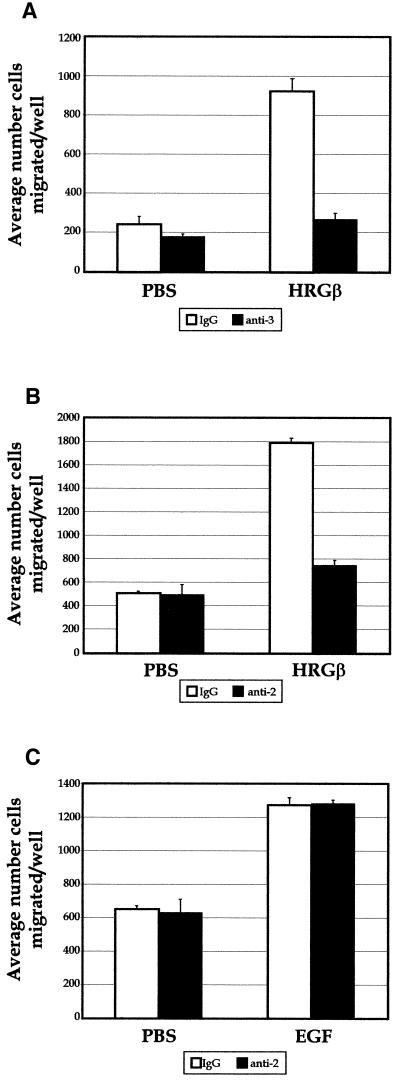
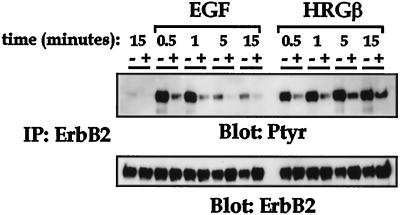
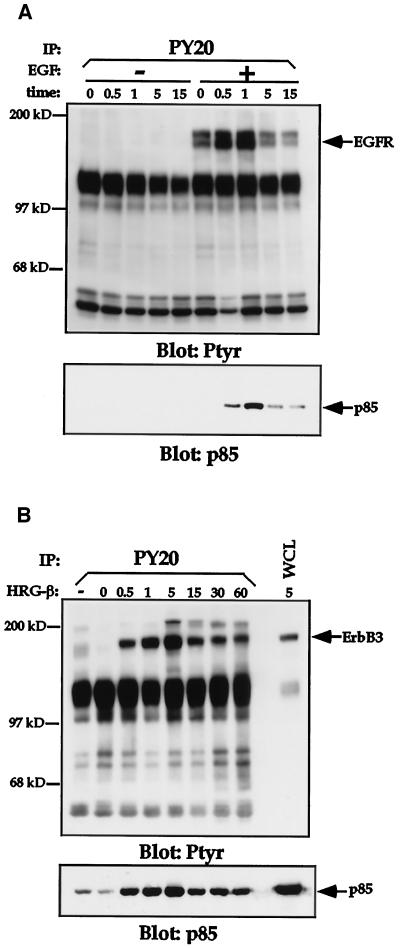
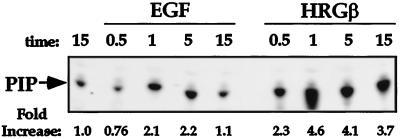
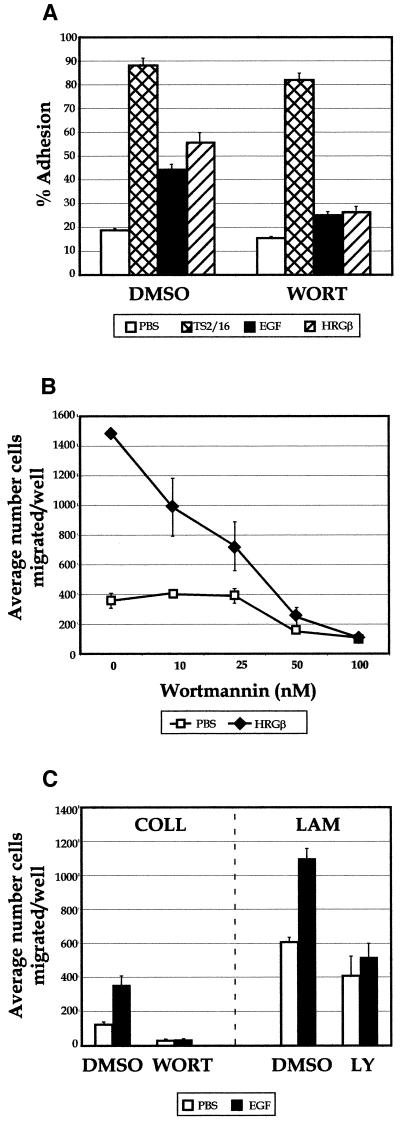
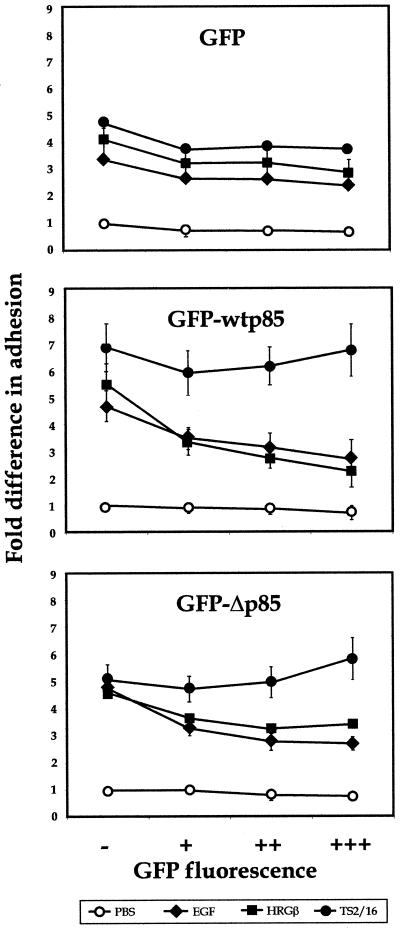
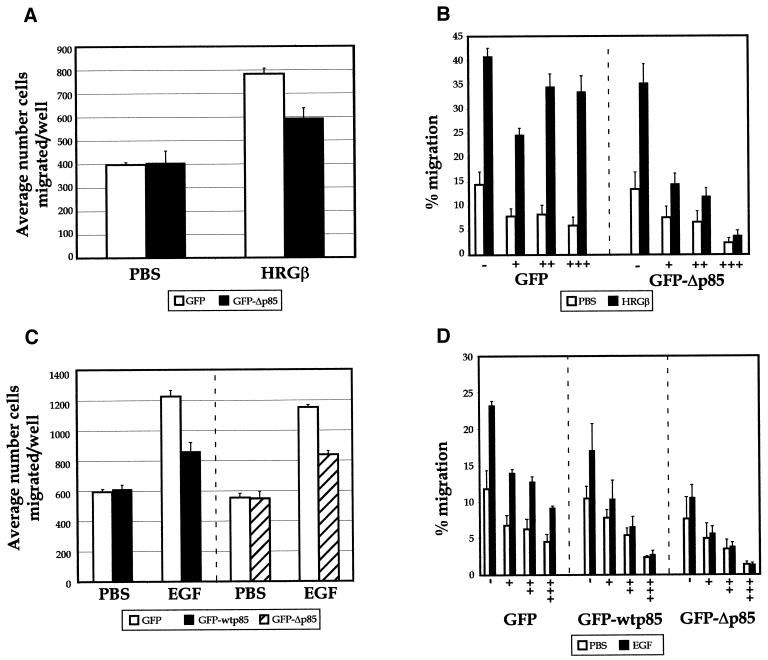
Integrins and growth factor receptors are important participants in cellular adhesion and migration. The EGF receptor (EGFR) family of tyrosine kinases and the beta1-integrin adhesion receptors are of particular interest, given the implication for their involvement in the initiation and progression of tumorigenesis. We used adhesion and chemotaxis assays to further elucidate the relationship between these two families of transmembrane signaling molecules. Specifically, we examined integrin-mediated adhesive and migratory characteristics of the metastatic breast carcinoma cell line MDA-MB-435 in response to stimulation with growth factors that bind to and activate the EGFR or erbB3 in these cells. Although ligand engagement of the EGFR stimulated modest beta1-dependent increases in cell adhesion and motility, heregulin-beta (HRGbeta) binding to the erbB3 receptor initiated rapid and potent induction of breast carcinoma cell adhesion and migration and required dimerization of erbB3 with erbB2. Pharmacologic inhibitors of phosphoinositide 3-OH kinase (PI 3-K) or transient expression of dominant negative forms of PI 3-K inhibited both EGF- and HRGbeta-mediated adhesion and potently blocked HRGbeta- and EGF-induced cell motility. Our results illustrate the critical role of PI 3-K activity in signaling pathways initiated by the EGFR or erbB3 to up-regulate beta1-integrin function.
Figures
References
-
- Adam L, Vadlamudi R, Kondapaka SB, Chernoff J, Mendelsohn J, Kumar R. Heregulin regulates cytoskeletal reorganization and cell migration through the p21-activated kinase-1 via phosphatidylinositol-3 kinase. J Biol Chem. 1998;273:28238–28246. - PubMed
-
- Akiyama SK, Larjava H, Yamada KM. Differences in the biosynthesis and localization of the fibronectin receptor in normal and transformed cultured human cells. Cancer Res. 1990;50:1601–1607. - PubMed
-
- Albelda SM. Role of integrins and other cell adhesion molecules in tumor progression and metastasis. Lab Invest. 1993;68:4–17. - PubMed
-
- Alroy I, Yarden Y. The ErbB signaling network in embryogenesis and oncogenesis: signal diversification through combinatorial ligand-receptor interactions. FEBS Lett. 1997;410:83–86. - PubMed
-
- Aplin AE, Howe A, Alahari SK, Juliani RL. Signal transduction and signal modulation by cell adhesion receptors: the role of integrins, cadherins, immunoglobulin-cell adhesion molecules, and selectins. Pharmacol Rev. 1998;50:197–263. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
