

Biparental inheritance of gamma-tubulin during human fertilization: molecular reconstitution of functional zygotic centrosomes in inseminated human oocytes and in cell-free extracts nucleated by human sperm
- PMID: 10473639
- PMCID: PMC25540
- DOI: 10.1091/mbc.10.9.2955
Biparental inheritance of gamma-tubulin during human fertilization: molecular reconstitution of functional zygotic centrosomes in inseminated human oocytes and in cell-free extracts nucleated by human sperm
Abstract
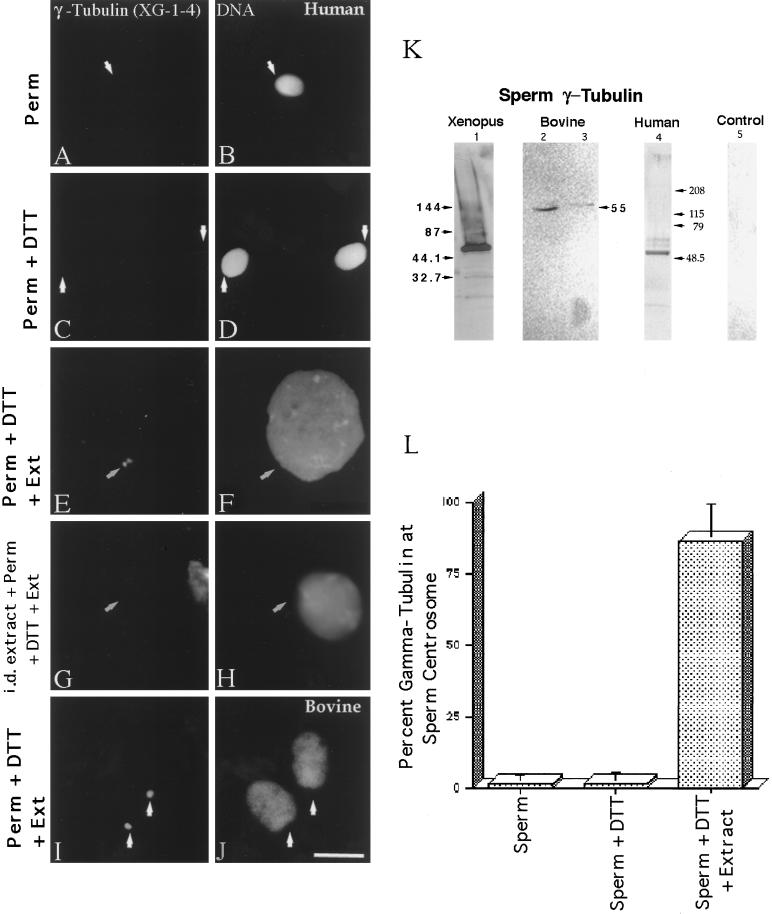
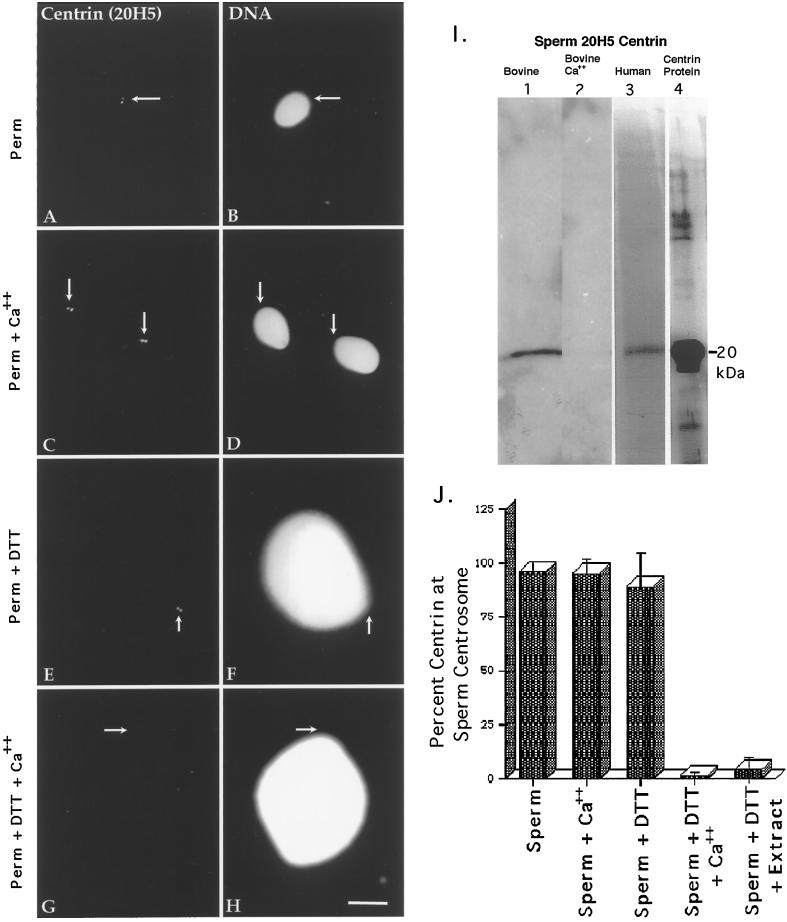
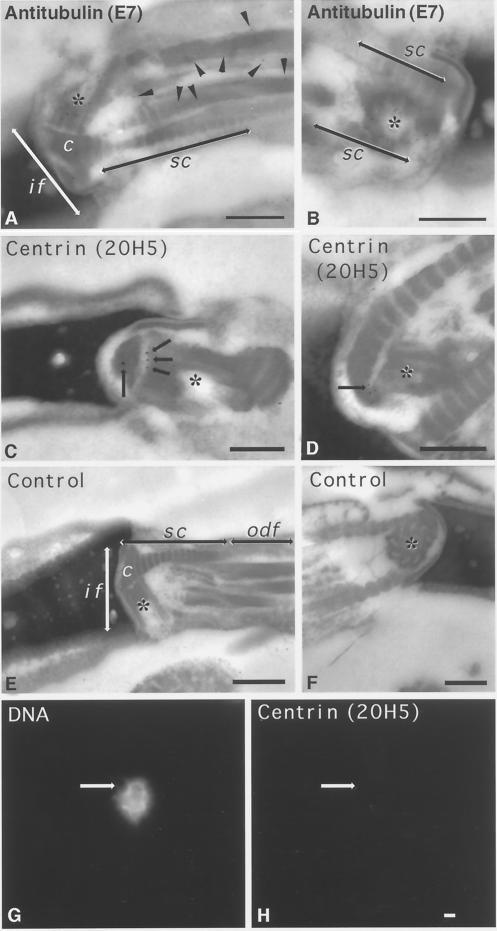
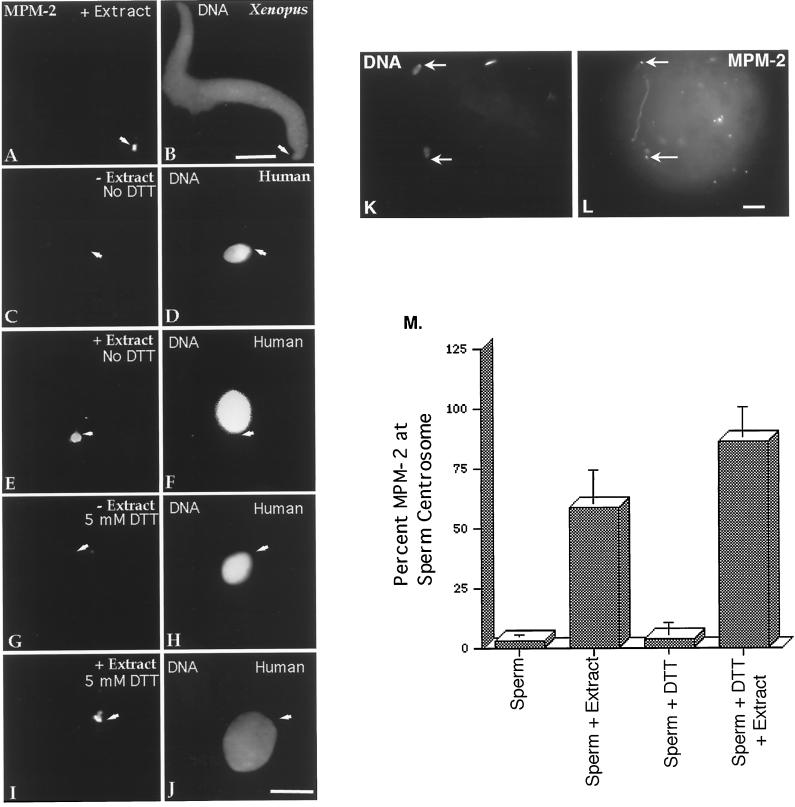
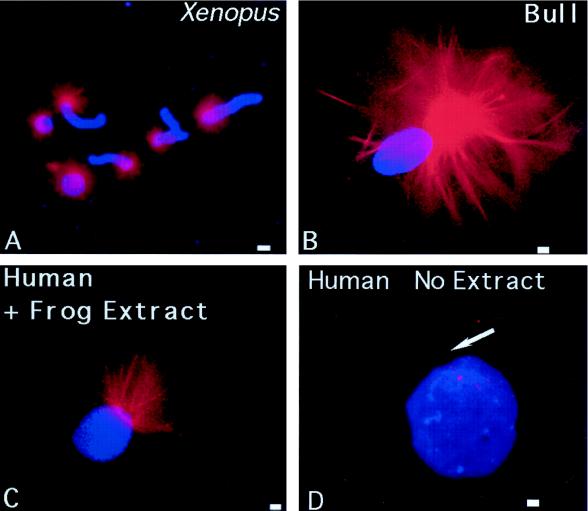
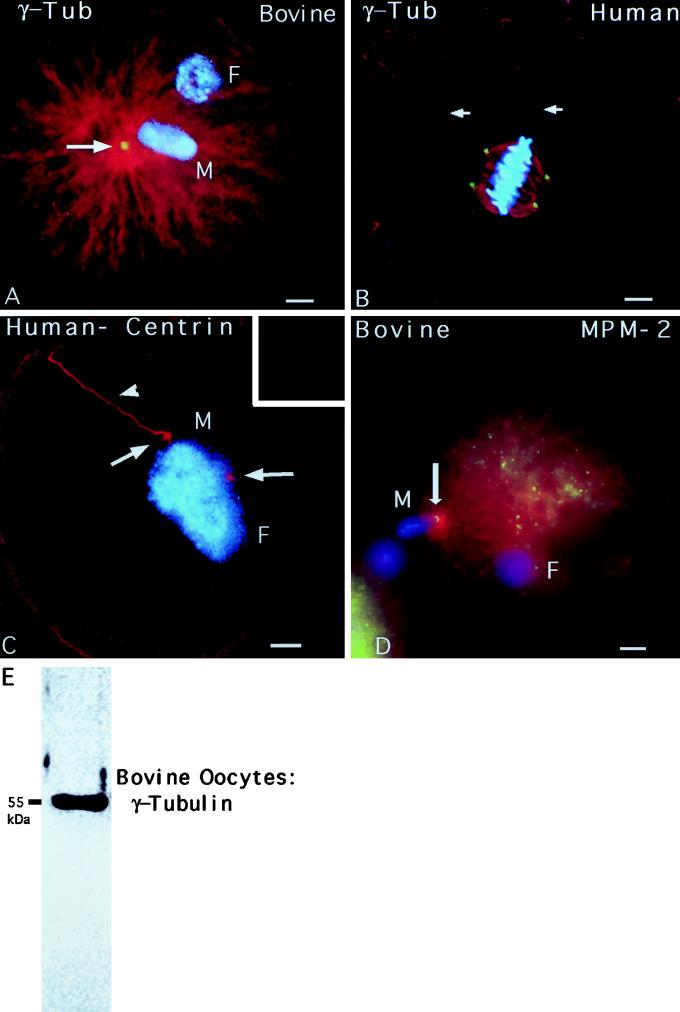
Human sperm centrosome reconstitution and the parental contributions to the zygotic centrosome are examined in mammalian zygotes and after exposure of spermatozoa to Xenopus laevis cell-free extracts. The presence and inheritance of the conserved centrosomal constituents gamma-tubulin, centrin, and MPM-2 (which detects phosphorylated epitopes) are traced, as is the sperm microtubule-nucleating capability on reconstituted centrosomes. gamma-Tubulin is biparentally inherited in humans (maternal >> than paternal): Western blots detect the presence of paternal gamma-tubulin. Recruitment of maternal gamma-tubulin to the sperm centrosome occurs after sperm incorporation in vivo or exposure to cell-free extract, especially after sperm "priming" induced by disulfide bond reduction. Centrin is found in the proximal sperm centrosomal region, demonstrates expected calcium sensitivity, but appears absent from the zygotic centrosome after sperm incorporation or exposure to extracts. Sperm centrosome phosphorylation is detected after exposure of primed sperm to egg extracts as well as during the early stages of sperm incorporation after fertilization. Finally, centrosome reconstitution in cell-free extracts permits sperm aster microtubule assembly in vitro. Collectively, these results support a model of a blended zygotic centrosome composed of maternal constituents attracted to an introduced paternal template after insemination.
Figures
Similar articles
-
Centrosome assembly in vitro: role of gamma-tubulin recruitment in Xenopus sperm aster formation.J Cell Biol. 1994 Jan;124(1-2):19-31. doi: 10.1083/jcb.124.1.19. J Cell Biol. 1994. PMID: 8294501 Free PMC article.
-
In vitro reconstitution of centrosome assembly and function: the central role of gamma-tubulin.Cell. 1994 Feb 25;76(4):623-37. doi: 10.1016/0092-8674(94)90503-7. Cell. 1994. PMID: 8124706
-
The centrosome and its mode of inheritance: the reduction of the centrosome during gametogenesis and its restoration during fertilization.Dev Biol. 1994 Oct;165(2):299-335. doi: 10.1006/dbio.1994.1256. Dev Biol. 1994. PMID: 7958403 Review.
-
Functional assessment of centrosomes of spermatozoa and spermatids microinjected into rabbit oocytes.Mol Reprod Dev. 2009 Mar;76(3):270-7. doi: 10.1002/mrd.20951. Mol Reprod Dev. 2009. PMID: 18646048
-
Characterization of human gamete centrosomes for assisted reproduction.Ital J Anat Embryol. 2001;106(2 Suppl 2):61-73. Ital J Anat Embryol. 2001. PMID: 11732597 Review.
Cited by
-
Centriolar defects, centrin 1 alterations, and FISH studies in human spermatozoa of a male partner of a couple that produces aneuploid embryos in natural and artificial fertilization.J Assist Reprod Genet. 2021 May;38(5):1197-1205. doi: 10.1007/s10815-021-02109-0. Epub 2021 Feb 22. J Assist Reprod Genet. 2021. PMID: 33619679 Free PMC article.
-
Functional Analysis of Human Pathological Semen Samples in an Oocyte Cytoplasmic Ex Vivo System.Sci Rep. 2018 Oct 18;8(1):15348. doi: 10.1038/s41598-018-33468-x. Sci Rep. 2018. PMID: 30337543 Free PMC article.
-
Poor centrosomal function of cat testicular spermatozoa impairs embryo development in vitro after intracytoplasmic sperm injection.Biol Reprod. 2006 Aug;75(2):252-60. doi: 10.1095/biolreprod.106.051342. Epub 2006 May 10. Biol Reprod. 2006. PMID: 16687647 Free PMC article.
-
The Role of Sperm Centrioles in Human Reproduction - The Known and the Unknown.Front Cell Dev Biol. 2019 Oct 1;7:188. doi: 10.3389/fcell.2019.00188. eCollection 2019. Front Cell Dev Biol. 2019. PMID: 31632960 Free PMC article. Review.
-
A novel atypical sperm centriole is functional during human fertilization.Nat Commun. 2018 Jun 7;9(1):2210. doi: 10.1038/s41467-018-04678-8. Nat Commun. 2018. PMID: 29880810 Free PMC article.
References
-
- Baron AT, Greenwood TM, Bazinet CW, Salisbury JL. Centrin is a component of the pericentriolar lattice. Biol Cell. 1992;76:383–388. - PubMed
-
- Baron AT, Suman VJ, Nemeth E, Salisbury JL. The pericentriolar lattice of PtK2 cells exhibits temperature and calcium-modulated behavior. J Cell Sci. 1994;107:2993–3003. - PubMed
-
- Bellvé AR, Zheng W, Martinova YS. Recovery, capacitation, acrosome reaction, and fractionation of sperm. Methods Enzymol. 1993;225:113–136. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
